Welcome to the Nanopore Community
Order MinION devices and consumables
Visit vwr.comPCR tiling of SARS-CoV-2 virus - classic protocol (SQK-LSK109 with EXP-NBD196)
Version for device: MinION
Introduction to the protocol
Overview of the protocol
-
Introduction to the protocol
To enable support for the rapidly expanding user requests, the team at Oxford Nanopore Technologies have put together an end-to-end workflow based on the ARTIC Network protocols and analysis methods.
While this protocol is available in the Nanopore Community, we kindly ask users to ensure they are citing the members of the ARTIC network who have been behind the development of these methods.
This protocol is based on the ARTIC amplicon sequencing protocol for MinION for nCOV 2019 by Josh Quick. The protocol generates 400 bp amplicons in a tiled fashion across the whole SARS-CoV-2 genome. Some example data is shown in the Downstream analysis and expected results section, this is generated using human coronavirus 229E to show what would be expected when running this protocol with SARS-CoV-2 samples.
Primers were designed by Josh Quick using Primal Scheme; the primer sequences can be found here.
Steps in the sequencing workflow:
Prepare for your experiment
you will need to:- Extract your RNA
- Ensure you have your sequencing kit, the correct equipment and third-party reagents
- Download the software for acquiring and analysing your data
- Check your flow cell to ensure it has enough pores for a good sequencing run
Prepare your library
You will need to:- Reverse transcribe your RNA samples with random hexamers
- Amplify the samples by tiled PCR using separate primer pools
- Combine the primer pools, purify and quantify the PCR products
- Prepare the DNA ends for adapter attachment
- Ligate native barcodes supplied in the kit to the DNA ends and pool the samples
- Ligate the sequencing adapters supplied in the kit to the DNA ends
- Prime the flow cell and load your DNA library into the flow cell

Sequencing and analysis
You will need to:- Start a sequencing run using the MinKNOW software, which will collect raw data from the device and convert it into basecalled reads
- Start the EPI2ME software and select the barcoding workflow
-
Before starting
This protocol outlines how to carryout PCR tiling of SARS-CoV-2 viral RNA samples on a 96-well plate using the Native Barcoding Expansion 96 (EXP-NBD196).
It is required to use total RNA extracted from samples that have been screened by a suitable qPCR assay. Here, we demonstrate the level of sensitivity and specificity by titrating total RNA extracted from cell culture infected with Human coronavirus 229E spiked into 100 ng human RNA extracted from GM12878 to give approximate figures.
Although not tested here, work performed by Josh Quick et al. on the Zika virus gives approximate dilution factors that may help the reduction of inhibiting compounds that can be co-extracted from samples.
Note: this is a guideline and not currently tested for COVID-19.
qPCR ct Dilution factor 18–35 none 15–18 1:10 12–15 1:100 When processing multiple samples at once, we recommend making master mixes with an additional 10% of the volume. We also recommend using pre- and post-PCR hoods when handling master mixes and samples. It is important to clean and/or UV irradiate these hoods between sample batches. Furthermore, to track and monitor cross-contamination events, it is important to run a negative control reaction at the reverse transcription stage using nuclease-free water instead of sample, and carrying this control through the rest of the prep.
To minimise the chance of pipetting errors when preparing primer mixes, we recommend ordering the tiling primers from IDT in a lab-ready format at 100 µM.
-
Kits in batches NBD196.10.0007 onwards have barcodes ordered in columns on the plate:

Kits in batches prior to NBD196.10.0007 have barcodes ordered in rows:




Equipment and consumables
- Materials
-
- Input RNA in 10 mM Tris-HCl, pH 8.0
- Ligation Sequencing Kit (SQK-LSK109)
- Flow Cell Priming Kit (EXP-FLP002)
- Native Barcoding Expansion 96 (EXP-NBD196)
- SFB Expansion (EXP-SFB001)
- Adapter Mix II Expansion (EXP-AMII001)
- Consumables
-
- LunaScript™ RT SuperMix Kit (NEB, cat # E3010)
- Q5® Hot Start High-Fidelity 2X Master Mix (NEB, cat # M0494)
- SARS-CoV-2 primers (lab-ready at 100 µM, IDT)
- Nuclease-free water (e.g. ThermoFisher, AM9937)
- Agencourt AMPure XP beads (Beckman Coulter™, A63881)
- Freshly prepared 80% ethanol in nuclease-free water
- Qubit dsDNA HS Assay Kit (ThermoFisher, Q32851)
- NEB Blunt/TA Ligase Master Mix (NEB, M0367)
- NEBNext® Ultra II End Repair / dA-tailing Module (NEB, E7546)
- NEBNext Quick Ligation Module (NEB, E6056)
- DNA 12000 Kit & Reagents - optional (Agilent Technologies)
- 1.5 ml Eppendorf DNA LoBind tubes
- Eppendorf twin.tec® PCR plate 96 LoBind, semi-skirted (Eppendorf™, cat # 0030129504) with heat seals
- 96-well deepwell plates
- 5 ml centrifuge tubes
- Equipment
-
- Hula mixer (gentle rotator mixer)
- Magnetic rack suitable for 96-well PCR plates, e.g. DynaMag™-96 Side Skirted Magnet (Thermo Fisher, cat # 12027)
- Microplate centrifuge, e.g. Fisherbrand™ Mini Plate Spinner Centrifuge (Fisher Scientific, 11766427)
- Vortex mixer
- Thermal cycler
- Stepper pipette and tips
- P1000 pipette and tips
- P200 pipette and tips
- P100 pipette and tips
- P20 pipette and tips
- P10 pipette and tips
- P2 pipette and tips
- Ice bucket with ice
- Timer
- Optional Equipment
-
- Agilent Bioanalyzer (or equivalent)
- Qubit fluorometer (or equivalent for QC check)
- Eppendorf 5424 centrifuge (or equivalent)
- PCR hood with UV steriliser (optional but recommended to reduce cross-contamination)
- PCR-Cooler (Eppendorf)
-
Input RNA guidelines
Where sample RNA is added to the below reaction, it is likely advantageous to follow the dilution guidelines proposed by Josh Quick:
qPCR Ct Dilution factor 18–35 none 15–18 1:10 12–15 1:100 If the sample has a low copy number (ct 18–35), use up to 16 µl of sample. Use nuclease-free water to make up any remaining volume. Take note to be aware that co-extracted compounds may inhibit reverse transcription and PCR.
-
Ligation Sequencing Kit contents (SQK-LSK109)

Name Acronym Cap colour No. of vials Fill volume per vial (µl) DNA CS DCS Yellow 1 50 Adapter Mix AMX Green 1 40 Ligation Buffer LNB Clear 1 200 L Fragment Buffer LFB White cap, orange stripe on label 2 1,800 S Fragment Buffer SFB Grey 2 1,800 Sequencing Buffer SQB Red 2 300 Elution Buffer EB Black 1 200 Loading Beads LB Pink 1 360 -
Flow Cell Priming Kit contents (EXP-FLP002)
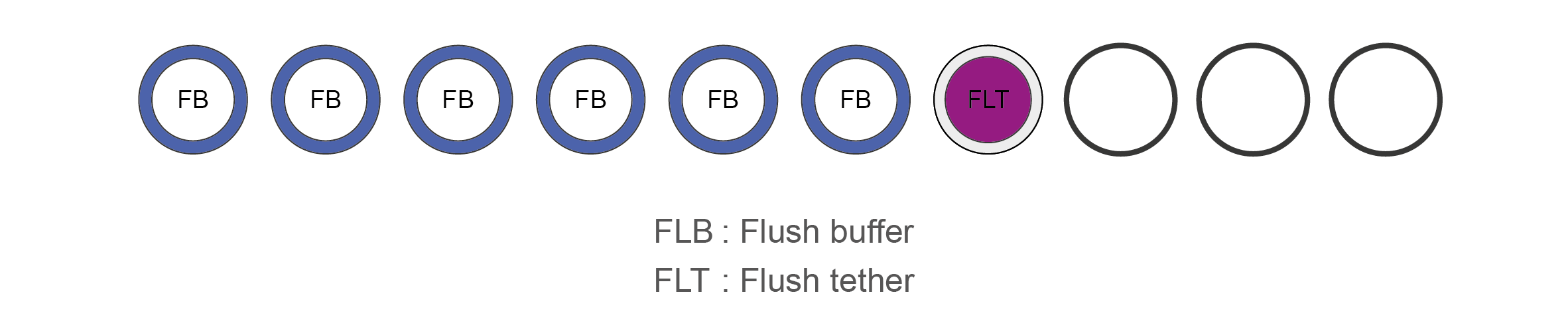
Name Acronym Cap colour No. of vials Fill volume per vial (μl) Flush Buffer FB Blue 6 1,170 Flush Tether FLT Purple 1 200 -
Native Barcoding Expansion 96 (EXP-NBD196) contents
Kits in batches NBD196.10.0007 onwards have barcodes ordered in columns on the plate:
Kits in batches prior to NBD196.10.0007 have barcodes ordered in rows:
Name Acronym Cap colour No. of vials Fill volume per vial (μl) Native Barcode 01-96 NB01-96 - 1 plate 40 μl per well Adapter Mix II AMII Green 1 70 -
SFB Expansion contents (EXP-SFB001)

Name Acronym Cap colour No. of vials Fill volume per vial (μl) Short Fragment Buffer SFB Grey 4 1,800 -
Adapter Mix II Expansion contents (EXP-AMII001)

Name Acronym Cap colour No. of tubes Fill volume per vial (μl) Adapter Mix II AMII Green 2 40 -
Adapter Mix II Expansion use
Protocols that use the Native Barcoding Expansions require 5 μl of AMII per reaction. Native Barcoding Expansions EXP-NBD104/NBD114 and EXP-NBD196 contain sufficient AMII for 6 and 12 reactions, respectively (or 12 and 24 reactions when sequencing on Flongle). This assumes that all barcodes are used in one sequencing run.
The Adapter Mix II expansion provides additional AMII for customers who are running subsets of barcodes, and allows a further 12 reactions (24 on Flongle).
-
Native barcode sequences
Component Forward sequence Reverse sequence NB01 CACAAAGACACCGACAACTTTCTT AAGAAAGTTGTCGGTGTCTTTGTG NB02 ACAGACGACTACAAACGGAATCGA TCGATTCCGTTTGTAGTCGTCTGT NB03 CCTGGTAACTGGGACACAAGACTC GAGTCTTGTGTCCCAGTTACCAGG NB04 TAGGGAAACACGATAGAATCCGAA TTCGGATTCTATCGTGTTTCCCTA NB05 AAGGTTACACAAACCCTGGACAAG CTTGTCCAGGGTTTGTGTAACCTT NB06 GACTACTTTCTGCCTTTGCGAGAA TTCTCGCAAAGGCAGAAAGTAGTC NB07 AAGGATTCATTCCCACGGTAACAC GTGTTACCGTGGGAATGAATCCTT NB08 ACGTAACTTGGTTTGTTCCCTGAA TTCAGGGAACAAACCAAGTTACGT NB09 AACCAAGACTCGCTGTGCCTAGTT AACTAGGCACAGCGAGTCTTGGTT NB10 GAGAGGACAAAGGTTTCAACGCTT AAGCGTTGAAACCTTTGTCCTCTC NB11 TCCATTCCCTCCGATAGATGAAAC GTTTCATCTATCGGAGGGAATGGA NB12 TCCGATTCTGCTTCTTTCTACCTG CAGGTAGAAAGAAGCAGAATCGGA NB13 AGAACGACTTCCATACTCGTGTGA TCACACGAGTATGGAAGTCGTTCT NB14 AACGAGTCTCTTGGGACCCATAGA TCTATGGGTCCCAAGAGACTCGTT NB15 AGGTCTACCTCGCTAACACCACTG CAGTGGTGTTAGCGAGGTAGACCT NB16 CGTCAACTGACAGTGGTTCGTACT AGTACGAACCACTGTCAGTTGACG NB17 ACCCTCCAGGAAAGTACCTCTGAT ATCAGAGGTACTTTCCTGGAGGGT NB18 CCAAACCCAACAACCTAGATAGGC GCCTATCTAGGTTGTTGGGTTTGG NB19 GTTCCTCGTGCAGTGTCAAGAGAT ATCTCTTGACACTGCACGAGGAAC NB20 TTGCGTCCTGTTACGAGAACTCAT ATGAGTTCTCGTAACAGGACGCAA NB21 GAGCCTCTCATTGTCCGTTCTCTA TAGAGAACGGACAATGAGAGGCTC NB22 ACCACTGCCATGTATCAAAGTACG CGTACTTTGATACATGGCAGTGGT NB23 CTTACTACCCAGTGAACCTCCTCG CGAGGAGGTTCACTGGGTAGTAAG NB24 GCATAGTTCTGCATGATGGGTTAG CTAACCCATCATGCAGAACTATGC NB25 GTAAGTTGGGTATGCAACGCAATG CATTGCGTTGCATACCCAACTTAC NB26 CATACAGCGACTACGCATTCTCAT ATGAGAATGCGTAGTCGCTGTATG NB27 CGACGGTTAGATTCACCTCTTACA TGTAAGAGGTGAATCTAACCGTCG NB28 TGAAACCTAAGAAGGCACCGTATC GATACGGTGCCTTCTTAGGTTTCA NB29 CTAGACACCTTGGGTTGACAGACC GGTCTGTCAACCCAAGGTGTCTAG NB30 TCAGTGAGGATCTACTTCGACCCA TGGGTCGAAGTAGATCCTCACTGA NB31 TGCGTACAGCAATCAGTTACATTG CAATGTAACTGATTGCTGTACGCA NB32 CCAGTAGAAGTCCGACAACGTCAT ATGACGTTGTCGGACTTCTACTGG NB33 CAGACTTGGTACGGTTGGGTAACT AGTTACCCAACCGTACCAAGTCTG NB34 GGACGAAGAACTCAAGTCAAAGGC GCCTTTGACTTGAGTTCTTCGTCC NB35 CTACTTACGAAGCTGAGGGACTGC GCAGTCCCTCAGCTTCGTAAGTAG NB36 ATGTCCCAGTTAGAGGAGGAAACA TGTTTCCTCCTCTAACTGGGACAT NB37 GCTTGCGATTGATGCTTAGTATCA TGATACTAAGCATCAATCGCAAGC NB38 ACCACAGGAGGACGATACAGAGAA TTCTCTGTATCGTCCTCCTGTGGT NB39 CCACAGTGTCAACTAGAGCCTCTC GAGAGGCTCTAGTTGACACTGTGG NB40 TAGTTTGGATGACCAAGGATAGCC GGCTATCCTTGGTCATCCAAACTA NB41 GGAGTTCGTCCAGAGAAGTACACG CGTGTACTTCTCTGGACGAACTCC NB42 CTACGTGTAAGGCATACCTGCCAG CTGGCAGGTATGCCTTACACGTAG NB43 CTTTCGTTGTTGACTCGACGGTAG CTACCGTCGAGTCAACAACGAAAG NB44 AGTAGAAAGGGTTCCTTCCCACTC GAGTGGGAAGGAACCCTTTCTACT NB45 GATCCAACAGAGATGCCTTCAGTG CACTGAAGGCATCTCTGTTGGATC NB46 GCTGTGTTCCACTTCATTCTCCTG CAGGAGAATGAAGTGGAACACAGC NB47 GTGCAACTTTCCCACAGGTAGTTC GAACTACCTGTGGGAAAGTTGCAC NB48 CATCTGGAACGTGGTACACCTGTA TACAGGTGTACCACGTTCCAGATG NB49 ACTGGTGCAGCTTTGAACATCTAG CTAGATGTTCAAAGCTGCACCAGT NB50 ATGGACTTTGGTAACTTCCTGCGT ACGCAGGAAGTTACCAAAGTCCAT NB51 GTTGAATGAGCCTACTGGGTCCTC GAGGACCCAGTAGGCTCATTCAAC NB52 TGAGAGACAAGATTGTTCGTGGAC GTCCACGAACAATCTTGTCTCTCA NB53 AGATTCAGACCGTCTCATGCAAAG CTTTGCATGAGACGGTCTGAATCT NB54 CAAGAGCTTTGACTAAGGAGCATG CATGCTCCTTAGTCAAAGCTCTTG NB55 TGGAAGATGAGACCCTGATCTACG CGTAGATCAGGGTCTCATCTTCCA NB56 TCACTACTCAACAGGTGGCATGAA TTCATGCCACCTGTTGAGTAGTGA NB57 GCTAGGTCAATCTCCTTCGGAAGT ACTTCCGAAGGAGATTGACCTAGC NB58 CAGGTTACTCCTCCGTGAGTCTGA TCAGACTCACGGAGGAGTAACCTG NB59 TCAATCAAGAAGGGAAAGCAAGGT ACCTTGCTTTCCCTTCTTGATTGA NB60 CATGTTCAACCAAGGCTTCTATGG CCATAGAAGCCTTGGTTGAACATG NB61 AGAGGGTACTATGTGCCTCAGCAC GTGCTGAGGCACATAGTACCCTCT NB62 CACCCACACTTACTTCAGGACGTA TACGTCCTGAAGTAAGTGTGGGTG NB63 TTCTGAAGTTCCTGGGTCTTGAAC GTTCAAGACCCAGGAACTTCAGAA NB64 GACAGACACCGTTCATCGACTTTC GAAAGTCGATGAACGGTGTCTGTC NB65 TTCTCAGTCTTCCTCCAGACAAGG CCTTGTCTGGAGGAAGACTGAGAA NB66 CCGATCCTTGTGGCTTCTAACTTC GAAGTTAGAAGCCACAAGGATCGG NB67 GTTTGTCATACTCGTGTGCTCACC GGTGAGCACACGAGTATGACAAAC NB68 GAATCTAAGCAAACACGAAGGTGG CCACCTTCGTGTTTGCTTAGATTC NB69 TACAGTCCGAGCCTCATGTGATCT AGATCACATGAGGCTCGGACTGTA NB70 ACCGAGATCCTACGAATGGAGTGT ACACTCCATTCGTAGGATCTCGGT NB71 CCTGGGAGCATCAGGTAGTAACAG CTGTTACTACCTGATGCTCCCAGG NB72 TAGCTGACTGTCTTCCATACCGAC GTCGGTATGGAAGACAGTCAGCTA NB73 AAGAAACAGGATGACAGAACCCTC GAGGGTTCTGTCATCCTGTTTCTT NB74 TACAAGCATCCCAACACTTCCACT AGTGGAAGTGTTGGGATGCTTGTA NB75 GACCATTGTGATGAACCCTGTTGT ACAACAGGGTTCATCACAATGGTC NB76 ATGCTTGTTACATCAACCCTGGAC GTCCAGGGTTGATGTAACAAGCAT NB77 CGACCTGTTTCTCAGGGATACAAC GTTGTATCCCTGAGAAACAGGTCG NB78 AACAACCGAACCTTTGAATCAGAA TTCTGATTCAAAGGTTCGGTTGTT NB79 TCTCGGAGATAGTTCTCACTGCTG CAGCAGTGAGAACTATCTCCGAGA NB80 CGGATGAACATAGGATAGCGATTC GAATCGCTATCCTATGTTCATCCG NB81 CCTCATCTTGTGAAGTTGTTTCGG CCGAAACAACTTCACAAGATGAGG NB82 ACGGTATGTCGAGTTCCAGGACTA TAGTCCTGGAACTCGACATACCGT NB83 TGGCTTGATCTAGGTAAGGTCGAA TTCGACCTTACCTAGATCAAGCCA NB84 GTAGTGGACCTAGAACCTGTGCCA TGGCACAGGTTCTAGGTCCACTAC NB85 AACGGAGGAGTTAGTTGGATGATC GATCATCCAACTAACTCCTCCGTT NB86 AGGTGATCCCAACAAGCGTAAGTA TACTTACGCTTGTTGGGATCACCT NB87 TACATGCTCCTGTTGTTAGGGAGG CCTCCCTAACAACAGGAGCATGTA NB88 TCTTCTACTACCGATCCGAAGCAG CTGCTTCGGATCGGTAGTAGAAGA NB89 ACAGCATCAATGTTTGGCTAGTTG CAACTAGCCAAACATTGATGCTGT NB90 GATGTAGAGGGTACGGTTTGAGGC GCCTCAAACCGTACCCTCTACATC NB91 GGCTCCATAGGAACTCACGCTACT AGTAGCGTGAGTTCCTATGGAGCC NB92 TTGTGAGTGGAAAGATACAGGACC GGTCCTGTATCTTTCCACTCACAA NB93 AGTTTCCATCACTTCAGACTTGGG CCCAAGTCTGAAGTGATGGAAACT NB94 GATTGTCCTCAAACTGCCACCTAC GTAGGTGGCAGTTTGAGGACAATC NB95 CCTGTCTGGAAGAAGAATGGACTT AAGTCCATTCTTCTTCCAGACAGG NB96 CTGAACGGTCATAGAGTCCACCAT ATGGTGGACTCTATGACCGTTCAG




Computer requirements and software
Computer requirements and software
-
MinION Mk1B IT requirements
Sequencing on a MinION Mk1B requires a high-spec computer or laptop to keep up with the rate of data acquisition. For more information, refer to the MinION Mk1B IT requirements document.
-
Software for nanopore sequencing
MinKNOW
The MinKNOW software controls the nanopore sequencing device, collects sequencing data and basecalls in real time. You will be using MinKNOW for every sequencing experiment to sequence, basecall and demultiplex if your samples were barcoded.
For instructions on how to run the MinKNOW software, please refer to MinKNOW protocol.
-
Check your flow cell
We highly recommend that you check the number of pores in your flow cell prior to starting a sequencing experiment. This should be done within 12 weeks of purchasing for MinION/GridION/PromethION or within four weeks of purchasing Flongle Flow Cells. Oxford Nanopore Technologies will replace any flow cell with fewer than the number of pores in the table below, when the result is reported within two days of performing the flow cell check, and when the storage recommendations have been followed. To do the flow cell check, please follow the instructions in the Flow Cell Check document.
Flow cell Minimum number of active pores covered by warranty Flongle Flow Cell 50 MinION/GridION Flow Cell 800 PromethION Flow Cell 5000
Library preparation
Reverse transcription
Reverse transcription
- Materials
-
- Input RNA in 10 mM Tris-HCl, pH 8.0
- Consumables
-
- LunaScript™ RT SuperMix Kit (NEB, cat # E3010)
- 1.5 ml Eppendorf DNA LoBind tubes
- Eppendorf twin.tec® PCR plate 96 LoBind, semi-skirted (Eppendorf™, cat # 0030129504) with heat seals
- Equipment
-
- P200 pipette and tips
- P2 pipette and tips
- Thermal cycler
- Microfuge
- Ice bucket with ice
- Optional Equipment
-
- PCR-Cooler (Eppendorf)
- PCR hood with UV steriliser (optional but recommended to reduce cross-contamination)
-
Input RNA guidelines
Where sample RNA is added to the below reaction, it is likely advantageous to follow the dilution guidelines proposed by Josh Quick:
qPCR Ct Dilution factor 18–35 none 15–18 1:10 12–15 1:100 If the sample has a low copy number (ct 18–35), use up to 16 µl of sample. Use nuclease-free water to make up any remaining volume. Take note to be aware that co-extracted compounds may inhibit reverse transcription and PCR.
-
In a clean pre-PCR hood, mix together the following components in each well of a 96-well plate on ice or in a PCR cool rack, such as the Eppendorf PCR-Cooler:
Reagent Volume per well RNA sample 16 µl LunaScript RT SuperMix (5x) 4 µl Total 20 µl Note: We recommend using up to 16 µl of RNA sample. Use nuclease-free water to make up the final volume to 16 µl if required.
-
Mix gently by pipetting, and spin down. Return the plate to ice.
-
Preheat the thermal cycler to 25°C.
-
Incubate the samples in the thermal cycler using the following program:
Step Temperature Time Cycles Primer annealing 25°C 2 min 1 cDNA synthesis 55°C 10 min 1 Heat inactivation 95°C 1 min 1 Hold 4°C ∞
PCR and clean-up
PCR and clean-up
- Consumables
-
- SARS-CoV-2 primers (lab-ready at 100 µM, IDT)
- Q5® Hot Start High-Fidelity 2X Master Mix (NEB, cat # M0494)
- Nuclease-free water (e.g. ThermoFisher, AM9937)
- Agencourt AMPure XP beads (Beckman Coulter™, A63881)
- Freshly prepared 80% ethanol in nuclease-free water
- 1.5 ml Eppendorf DNA LoBind tubes
- Eppendorf twin.tec® PCR plate 96 LoBind, semi-skirted (Eppendorf™, cat # 0030129504) with heat seals
- 96-well deepwell plates
- 5 ml centrifuge tubes
- Equipment
-
- Microfuge
- Thermal cycler
- Hula mixer (gentle rotator mixer)
- Magnetic rack suitable for 96-well plates
- Multichannel pipette and tips
- P200 pipette and tips
- P100 pipette and tips
- P20 pipette and tips
- P10 pipette and tips
- P2 pipette and tips
- Optional Equipment
-
- Qubit fluorometer (or equivalent for QC check)
- Agilent Bioanalyzer (or equivalent)
- PCR hood with UV steriliser (optional but recommended to reduce cross-contamination)
-
Primer design
To generate tiled PCR amplicons from the SARS-CoV-2 viral cDNA, primers were designed by Josh Quick using Primal Scheme. These primers are designed to generate 400 bp amplicons that overlap by approximately 20 bp. These primer sequences can be found here. Where we show example data outputs in this protocol, the same parameters were used to design primers to the human coronavirus 229E to provide guideline statistics.
-
If primers are supplied individually, add 5 µl of each primer from pool A per sample to a 1.5 ml Eppendorf DNA LoBind tube to give a 100 µM stock primer pool.
-
If primers are supplied individually, add 5 µl of each primer from pool B per sample to a 1.5 ml Eppendorf DNA LoBind tube to give a 100 µM stock primer pool.
-
Dilute each 100 µM stock 1 in 10 with nuclease-free water to form a working stock of each pool at 10 µM.
Note: To achieve the desired final concentration of each primer in the pool at 0.015 µM in the PCR reaction, 3.7 µl of the 10 µM working stock is needed for each PCR reaction. Two separate PCR reactions will be performed per sample, one for pool A primers and one for pool B. This results in tiled amplicons that have approximately 20 bp overlap.
-
In a clean pre-PCR hood, set up two individual reactions using primer pool A and primer pool B per sample:
Reagent Volume (pool A) Volume (pool B) Q5® Hot Start High-Fidelity 2X Master Mix 12.5 µl 12.5 µl Primer pool at 10 µM (A or B) 3.7 µl 3.7 µl Nuclease-free water 3.8 µl 3.8 µl Total 20 µl 20 µl -
In a clean 96-well plate, aliquot 20 µl of pool A reaction to each well per sample. Repeat in a new 96-well plate with pool B reaction. Add 5 µl of the reverse-transcribed samples per well in both plates.
-
Mix well by pipetting and spin down in a centrifuge.
-
Incubate using the following program, with the heated lid set to 105°C:
Step Temperature Time Cycles Initial denaturation 98°C 30 sec 1 Denaturation
Annealing and extension98°C
65°C15 sec
5 min
25–35Hold 4°C ∞ Note: Cycle number should be varied for low or high viral load samples. Guidelines provided by Josh Quick suggest that 25 cycles should be used for Ct 18–21 up to a maximum of 35 cycles for Ct 35, however this has not been tested here.
-
Combine the 25 µl reaction from pool A and the 25 µl reaction from pool B per sample, into a new deep well plate; one well per sample.
Note: Ensure the sample from pool A corresponds to the sample from pool B.
-
Resuspend the AMPure XP beads by vortexing.
-
Add 50 µl of resuspended AMPure XP beads to each well and mix by gently pipetting.
-
Allow DNA to bind to the beads for 5 minutes at room temperature.
-
Prepare 50 ml of fresh 80% ethanol in nuclease-free water.
-
Spin down the 96-well plate and pellet the beads on a magnet for 5 minutes. Keep the plate on the magnet until the eluate is clear and colourless, and pipette off the supernatant.
-
Keep the plate on the magnet and wash the beads in each well with 200 µl of freshly prepared 80% ethanol without disturbing the pellet. Remove the ethanol using a pipette and discard.
-
Repeat the previous step.
-
Spin down and place the plate back on the magnet. Pipette off any residual ethanol. Allow to dry for ~ 30 seconds, but do not dry the pellet to the point of cracking.
-
Remove the plate from the magnetic rack and resuspend each pellet in 15 µl nuclease-free water. Incubate for 2 minutes at room temperature.
-
Pellet the beads on a magnet until the eluate is clear and colourless.
-
Remove and retain 15 µl of eluate containing the DNA library per well, into a clean 96-well plate.
Dispose of the pelleted beads.
-
Quantify 1 µl of each eluted sample using a Qubit fluorometer.
-
Store any unused amplified material at -20°C for use in later experiments.
-
Expected results
During initial method development, it is useful to analyse 1 µl on an Agilent Bioanalyzer chip or an appropriate amount on an agarose gel. The traces below show expected results where a dilution series of coronavirus 229E was spiked into 100 ng of human RNA extracted from GM12878 (primers were designed against the human coronavirus 229E reference genome using Primal Scheme). Here, Qubit hsDNA results and Agilent Bioanalyser (DNA 12000 assay) traces are shown for 30 and 35 cycles of PCR with input concentrations ranging from 10 pg to 0.001 pg in 100 ng human RNA. While not directly comparable to Ct values of a real biological sample, these give a rough approximation of high to low viral titres. A human-only and reverse transcription negative control were also included.
Note: The viral RNA that was used for this spike-in experiment was obtained from ATCC and is total RNA extracted from human cell lines infected with coronavirus 229E. So, 10 pg of spike-in represents a mix of human and viral RNA, spiked into 100 ng of human RNA extracted from GM12878 cells.
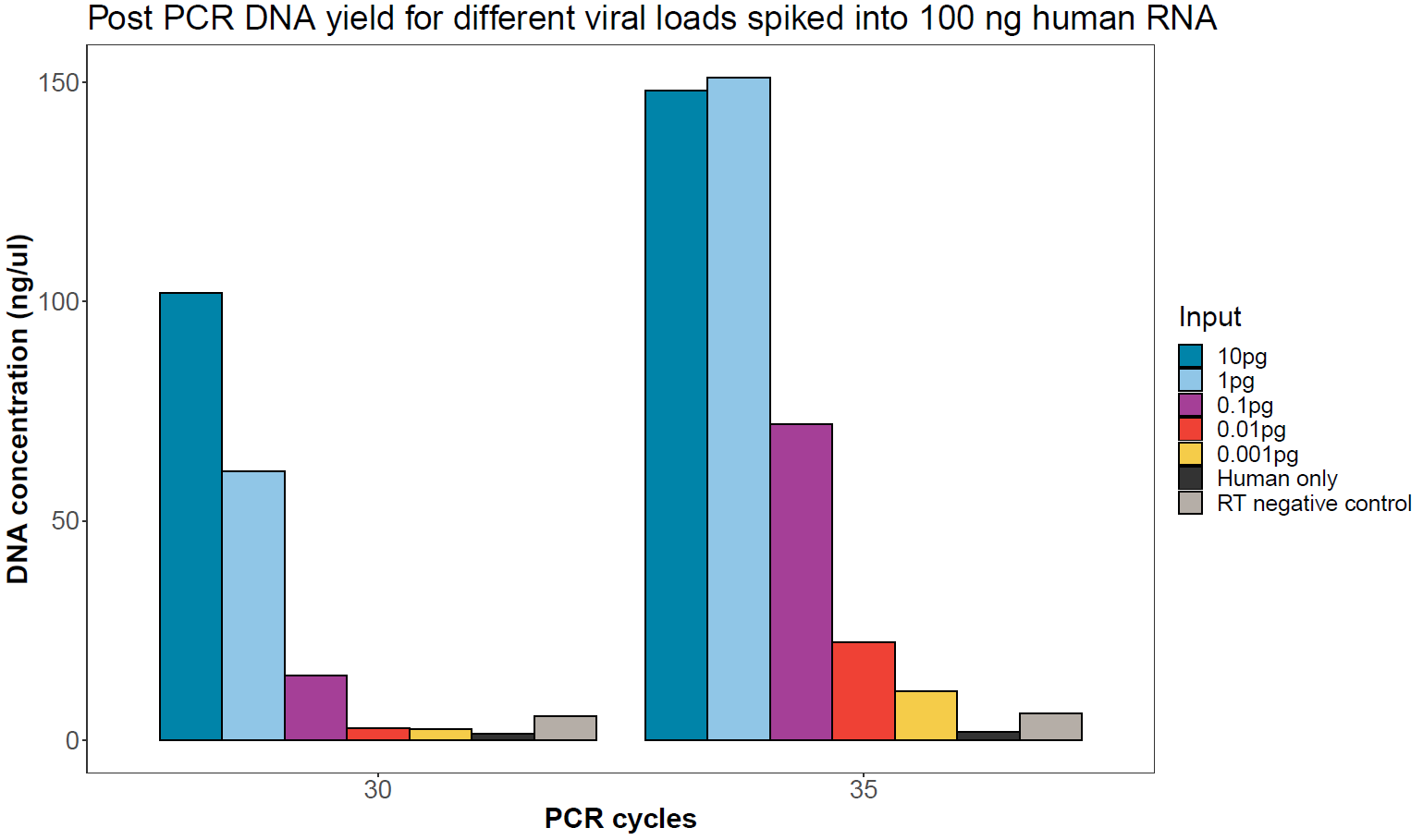
Figure 1. DNA yield after PCR and AMPure XP clean-up for decreasing viral input and different PCR cycle numbers in a background of 100 ng human RNA.For a 400 bp amplicon, approximately 50 ng (~0.2 pmol) is required for the end-prep step. PCR cycles can be adjusted based on initial results to minimise the number of cycles. For samples that provide less than 50 ng total yield, further PCRs may be carried out on the remaining reverse transcription reaction.
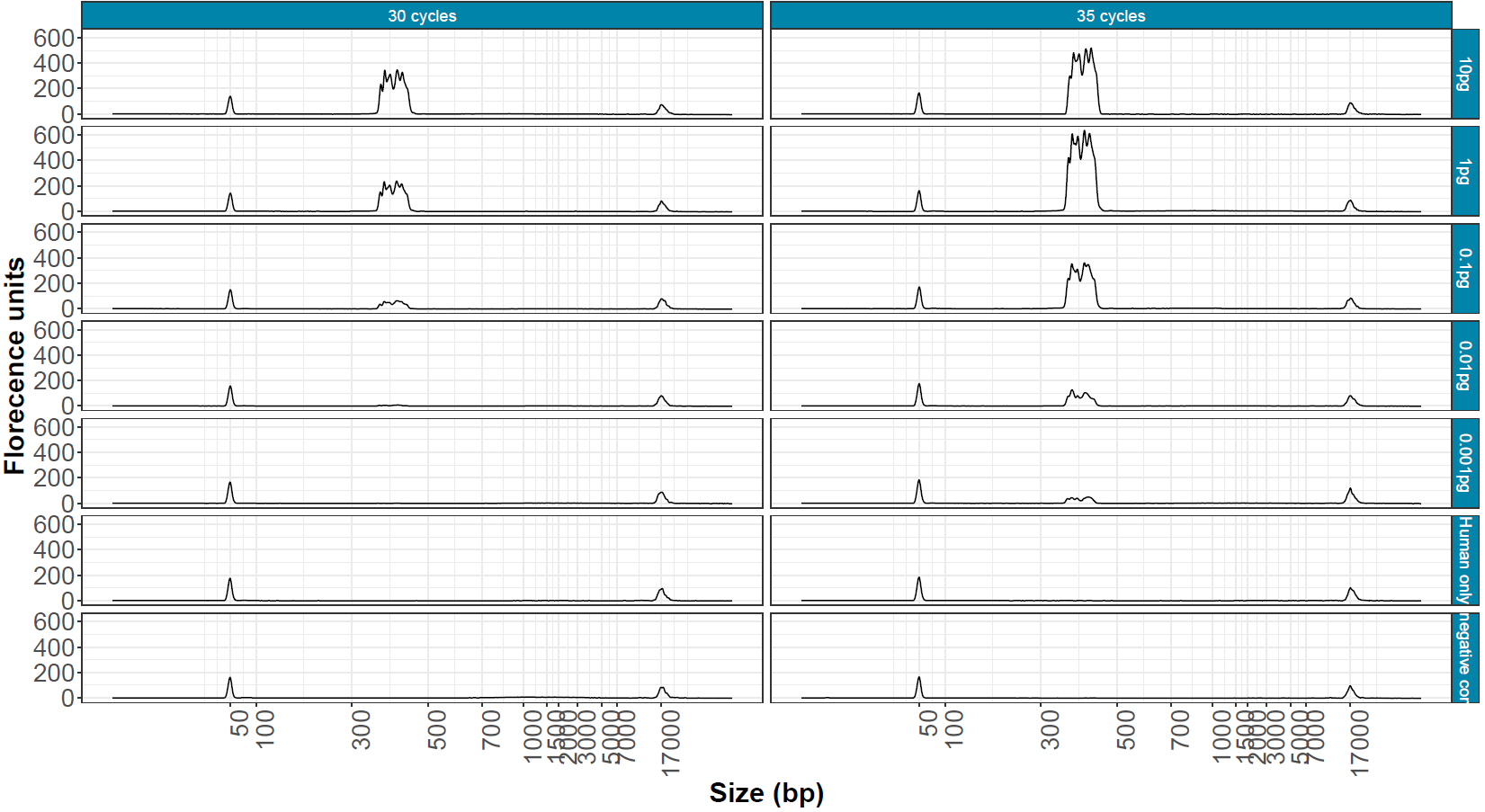
Figure 2. Bioanalyser traces of 1 µl of post-PCR cleaned up samples with decreasing input quantities, spiked into 100 ng human RNA amplified with 30 and 35 cycles. RT negative controls and human-only negative controls show no product in the 300–400 bp range.


End-prep
End-prep
- Consumables
-
- Nuclease-free water (e.g. ThermoFisher, AM9937)
- NEBNext® Ultra II End Repair / dA-tailing Module (NEB, E7546)
- 1.5 ml Eppendorf DNA LoBind tubes
- Eppendorf twin.tec® PCR plate 96 LoBind, semi-skirted (Eppendorf™, cat # 0030129504) with heat seals
- Equipment
-
- Multichannel pipette and tips
- P1000 pipette and tips
- P100 pipette and tips
- P10 pipette and tips
- Thermal cycler
- Microfuge
- Ice bucket with ice
-
Determine the volume of the cleaned-up PCR reaction that yields 200 fmol (50 ng) of DNA per sample and aliquot in a clean 96-well plate.
-
Prepare the NEBNext Ultra II End repair / dA-tailing Module reagents in accordance with manufacturer’s instructions, and place on ice.
For optimal performance, NEB recommend the following:
- Thaw all reagents on ice.
- Flick and/or invert reagent tube to ensure they are well mixed.
- Always spin down tubes before opening for the first time each day.
- The Ultra II End prep buffer and FFPE DNA Repair buffer may have a little precipitate. Allow the mixture to come to room temperature and pipette the buffer up and down several times to break up the precipitate, followed by vortexing the tube for several seconds to ensure the reagent is thoroughly mixed.
- The FFPE DNA repair buffer may have a yellow tinge and is fine to use if yellow.
-
Make up each sample per well to 12.5 µl using nuclease-free water.
-
Add the following components to each well:
Between each addition, pipette mix 10 - 20 times.
Reagent Volume Ultra II End-prep reaction buffer 1.75 µl Ultra II End-prep enzyme mix 0.75 µl Total 2.5 µl -
Mix well by pipetting and spin down in a centrifuge.
-
Using a thermal cycler, incubate at 20°C for 5 minutes and 65°C for 5 minutes.
Native barcode ligation
- Materials
-
- Native Barcoding Expansion 96 (EXP-NBD196)
- Short Fragment Buffer (SFB)
- Consumables
-
- Freshly prepared 80% ethanol in nuclease-free water
- 1.5 ml Eppendorf DNA LoBind tubes
- Nuclease-free water (e.g. ThermoFisher, AM9937)
- Agencourt AMPure XP beads (Beckman Coulter, A63881)
- NEB Blunt/TA Ligase Master Mix (NEB, M0367)
- Equipment
-
- Magnetic rack suitable for 96-well plates
- Thermal cycler
- Hula mixer (gentle rotator mixer)
- Vortex mixer
- Ice bucket with ice
- Microfuge
- P1000 pipette and tips
- P100 pipette and tips
- P10 pipette and tips
- Optional Equipment
-
- Qubit fluorometer (or equivalent for QC check)
-
Thaw the native barcodes at room temperature. Use one barcode per sample. Individually mix the barcodes by pipetting, spin down, and place them on ice.
-
Thaw the tube of Short Fragment Buffer (SFB) at room temperature, mix by vortexing, spin down and place on ice.
-
Select a unique barcode for every sample to be run.
-
In a new 96-well plate, add the reagents in the order given below per well:
Mix well by pipetting and spin down in a centrifuge.
Reagent Volume Nuclease-free water 3 µl End-prepped DNA 0.75 µl Native Barcode 1.25 µl Blunt/TA Ligase Master Mix 5 µl Total 10 µl -
Mix contents thoroughly by pipetting and spin down briefly.
-
Using a thermal cycler, incubate at 20°C for 20 mins and at 65°C for 10 mins.
-
Pool the barcoded library together and carry forward 480 µl of the library.
-
Resuspend the AMPure XP beads by vortexing.
-
Add 192 µl of resuspended AMPure XP beads to the 480 µl of pooled reaction and mix by pipetting.
-
Incubate on a Hula mixer (rotator mixer) for 10 minutes at room temperature.
-
Prepare 500 µl of fresh 80% ethanol in nuclease-free water.
-
Spin down the sample and pellet the beads on a magnet for 5 minutes. Keep the tube on the magnet until the eluate is clear and colourless, and pipette off the supernatant.
-
Wash the beads by adding 700 μl Short Fragment Buffer (SFB). Flick the beads to resuspend, then return the tube to the magnetic rack and allow the beads to pellet. Keep the tube on the magnet until the eluate is clear and colourless. Remove the supernatant using a pipette and discard.
-
Repeat the previous step.
-
Keep the tube on the magnet and wash the beads with 100 µl of freshly prepared 80% ethanol without disturbing the pellet. Remove the ethanol using a pipette and discard.
-
Spin down and place the tube back on the magnet. Pipette off any residual ethanol. Allow to dry for ~30 seconds, but do not dry the pellet to the point of cracking.
-
Remove the tube from the magnetic rack and resuspend the pellet in 35 µl nuclease-free water. Incubate for 2 minutes at room temperature.
-
Pellet the beads on a magnet until the eluate is clear and colourless.
-
Remove and retain 35 µl of eluate into a clean 1.5 ml Eppendorf DNA LoBind tube.
-
Quantify 1 µl of eluted sample using a Qubit fluorometer - recovery aim 2 ng/μl.
Adapter ligation and clean-up
Adapter ligation and clean-up
- Materials
-
- Elution Buffer (EB)
- Short Fragment Buffer (SFB)
- Adapter Mix II (AMII)
- Consumables
-
- NEBNext Quick Ligation Module (NEB, E6056)
- Agencourt AMPure XP beads (Beckman Coulter™, A63881)
- 1.5 ml Eppendorf DNA LoBind tubes
- Equipment
-
- Microfuge
- Magnetic rack
- Vortex mixer
- Hula mixer (gentle rotator mixer)
- Optional Equipment
-
- Qubit fluorometer (or equivalent for QC check)
-
Adapter Mix II Expansion use
Protocols that use the Native Barcoding Expansions require 5 μl of AMII per reaction. Native Barcoding Expansions EXP-NBD104/NBD114 and EXP-NBD196 contain sufficient AMII for 6 and 12 reactions, respectively (or 12 and 24 reactions when sequencing on Flongle). This assumes that all barcodes are used in one sequencing run.
The Adapter Mix II expansion provides additional AMII for customers who are running subsets of barcodes, and allows a further 12 reactions (24 on Flongle).
-
Thaw the Elution Buffer (EB), Short Fragment Buffer (SFB), and NEBNext Quick Ligation Reaction Buffer (5x) at room temperature and mix by vortexing. Then spin down and place on ice. Check the contents of each tube are clear of any precipitate.
-
Spin down the T4 Ligase and the Adapter Mix II (AMII), and place on ice.
-
Taking the pooled and barcoded DNA, perform adapter ligation as follows, mixing by flicking the tube between each sequential addition.
Reagent Volume Pooled barcoded sample (30-50 ng) 30 µl Adapter Mix II (AMII) 5 µl NEBNext Quick Ligation Reaction Buffer (5X) 10 µl Quick T4 DNA Ligase 5 µl Total 50 µl -
Ensure the components are thoroughly mixed by pipetting, and spin down.
-
Incubate the reaction for 10 minutes at room temperature.
-
Resuspend the AMPure XP beads by vortexing.
-
Add 20 µl of resuspended AMPure XP beads to the reaction and mix by pipetting.
-
Incubate on a Hula mixer (rotator mixer) for 10 minutes at room temperature.
-
Spin down the sample and pellet the beads on a magnet for 5 minutes. Keep the tube on the magnet until the eluate is clear and colourless, and pipette off the supernatant.
-
Wash the beads by adding 125 μl Short Fragment Buffer (SFB). Flick the beads to resuspend, then return the tube to the magnetic rack and allow the beads to pellet. Keep the tube on the magnet until the eluate is clear and colourless. Remove the supernatant using a pipette and discard.
-
Repeat the previous step.
-
Spin down and place the tube back on the magnet. Pipette off any residual supernatant. Allow to dry for ~30 seconds, but do not dry the pellet to the point of cracking.
-
Remove the tube from the magnetic rack and resuspend the pellet by pipetting in 15 µl Elution Buffer (EB). Spin down and incubate for 5 minutes at room temperature.
-
Pellet the beads on a magnet until the eluate is clear and colourless, for at least 1 minute.
-
Remove and retain 15 µl of eluate containing the DNA library into a clean 1.5 ml Eppendorf DNA LoBind tube.
Dispose of the pelleted beads
-
Quantify 1 µl of eluted sample using a Qubit fluorometer.
-
Optional ActionIf quantities allow, the library may be diluted in Elution Buffer (EB) for splitting across multiple flow cells.
Additional buffer for doing this can be found in the Sequencing Auxiliary Vials expansion (EXP-AUX001), available to purchase separately. This expansion also contains additional vials of Sequencing Buffer (SQB) and Loading Beads (LB), required for loading the libraries onto flow cells.
Priming and loading the SpotON flow cell
Priming and loading the SpotON flow cell
- Materials
-
- Flow Cell Priming Kit (EXP-FLP002)
- Loading Beads (LB)
- Sequencing Buffer (SQB)
- Consumables
-
- 1.5 ml Eppendorf DNA LoBind tubes
- Nuclease-free water (e.g. ThermoFisher, AM9937)
- Equipment
-
- MinION Mk1B or Mk1C
- SpotON Flow Cell
- P1000 pipette and tips
- P100 pipette and tips
- P20 pipette and tips
- P10 pipette and tips
-
Thaw the Sequencing Buffer (SQB), Loading Beads (LB), Flush Tether (FLT) and one tube of Flush Buffer (FB) at room temperature before mixing the reagents by vortexing, and spin down at room temperature.
-
To prepare the flow cell priming mix, add 30 µl of thawed and mixed Flush Tether (FLT) directly to the tube of thawed and mixed Flush Buffer (FB), and mix by vortexing at room temperature.
-
Open the MinION device lid and slide the flow cell under the clip.
Press down firmly on the flow cell to ensure correct thermal and electrical contact.


-
Optional ActionComplete a flow cell check to assess the number of pores available before loading the library.
This step can be omitted if the flow cell has been checked previously.
See the flow cell check instructions in the MinKNOW protocol for more information.
-
Slide the flow cell priming port cover clockwise to open the priming port.

-
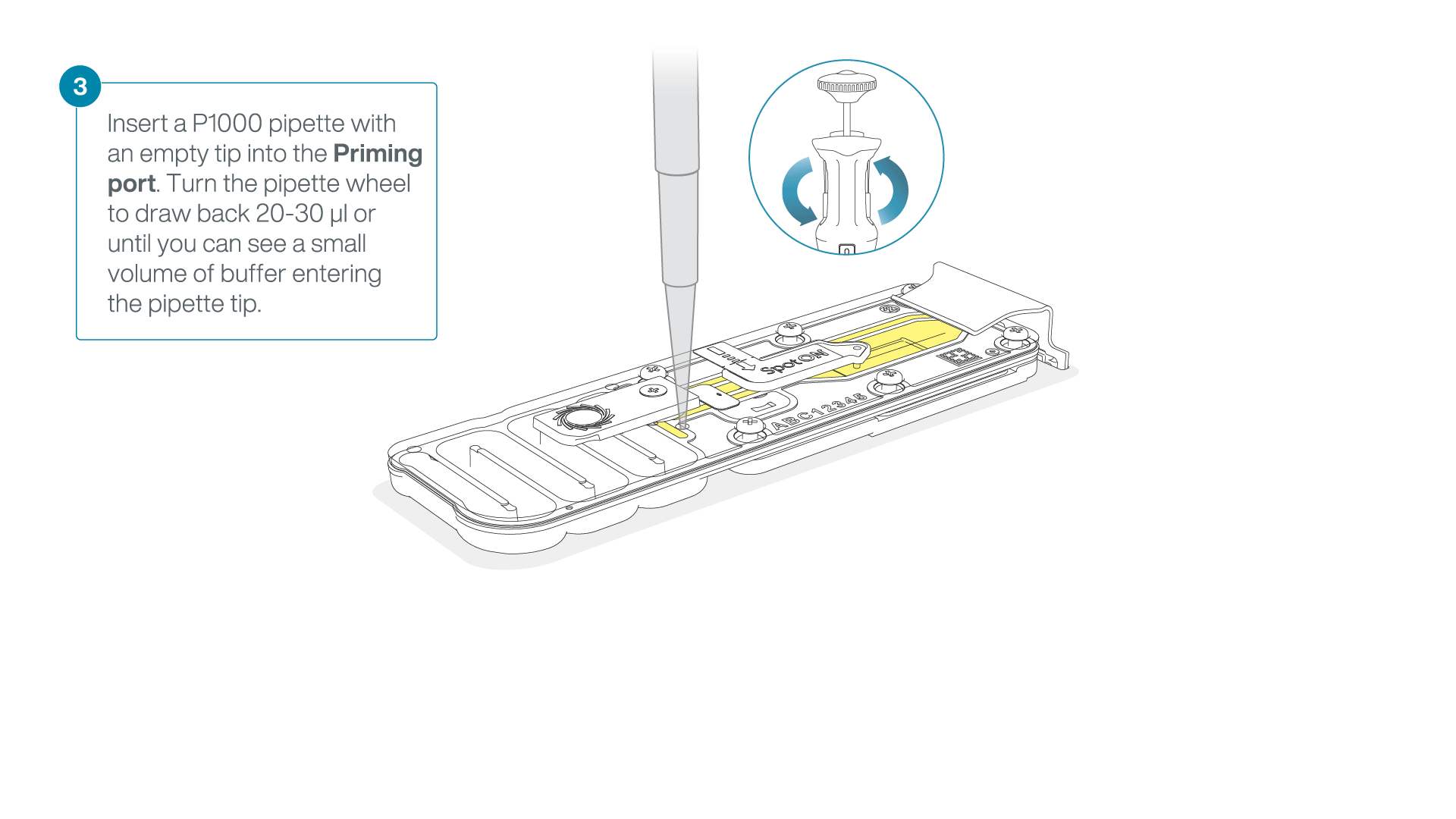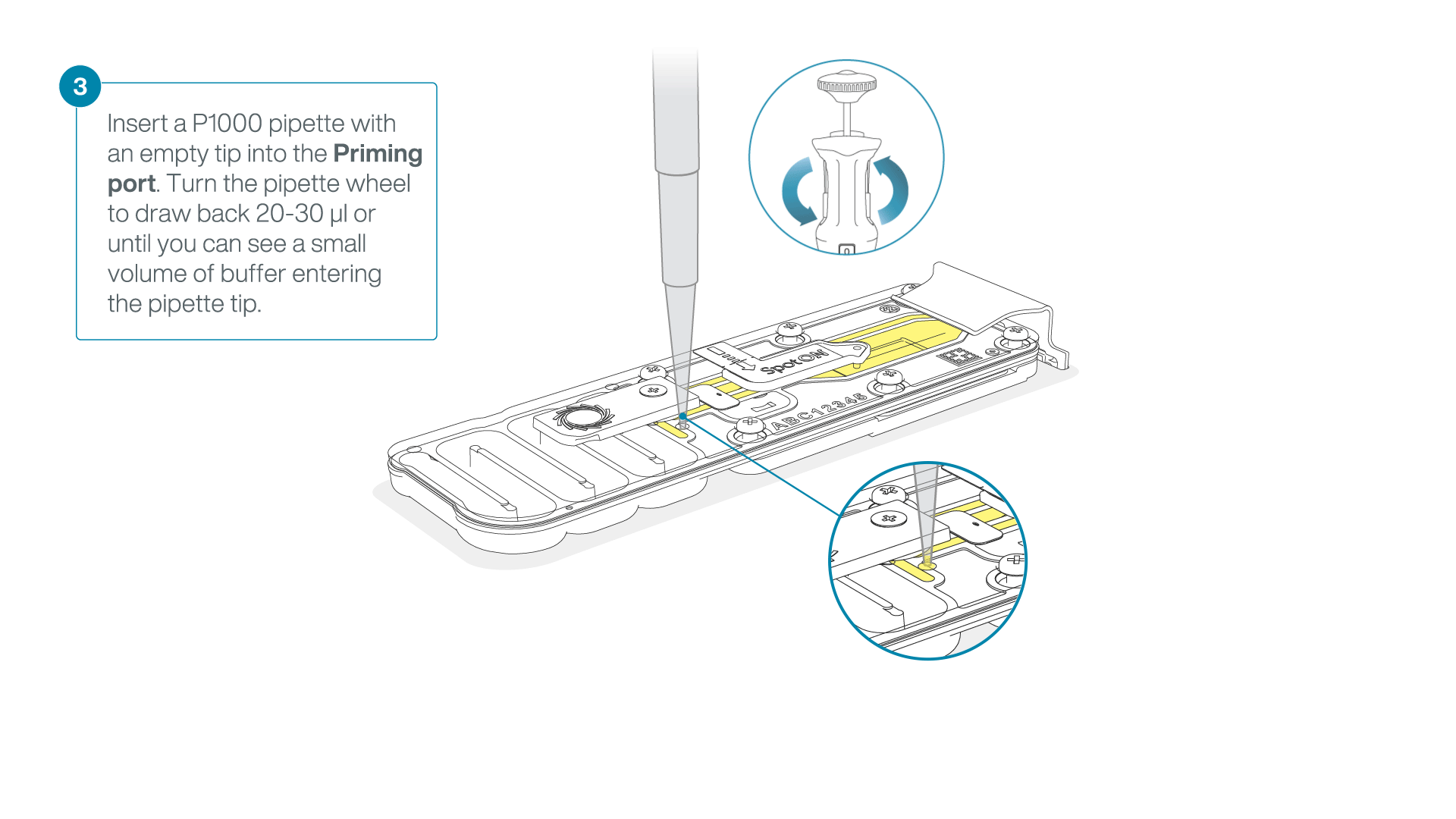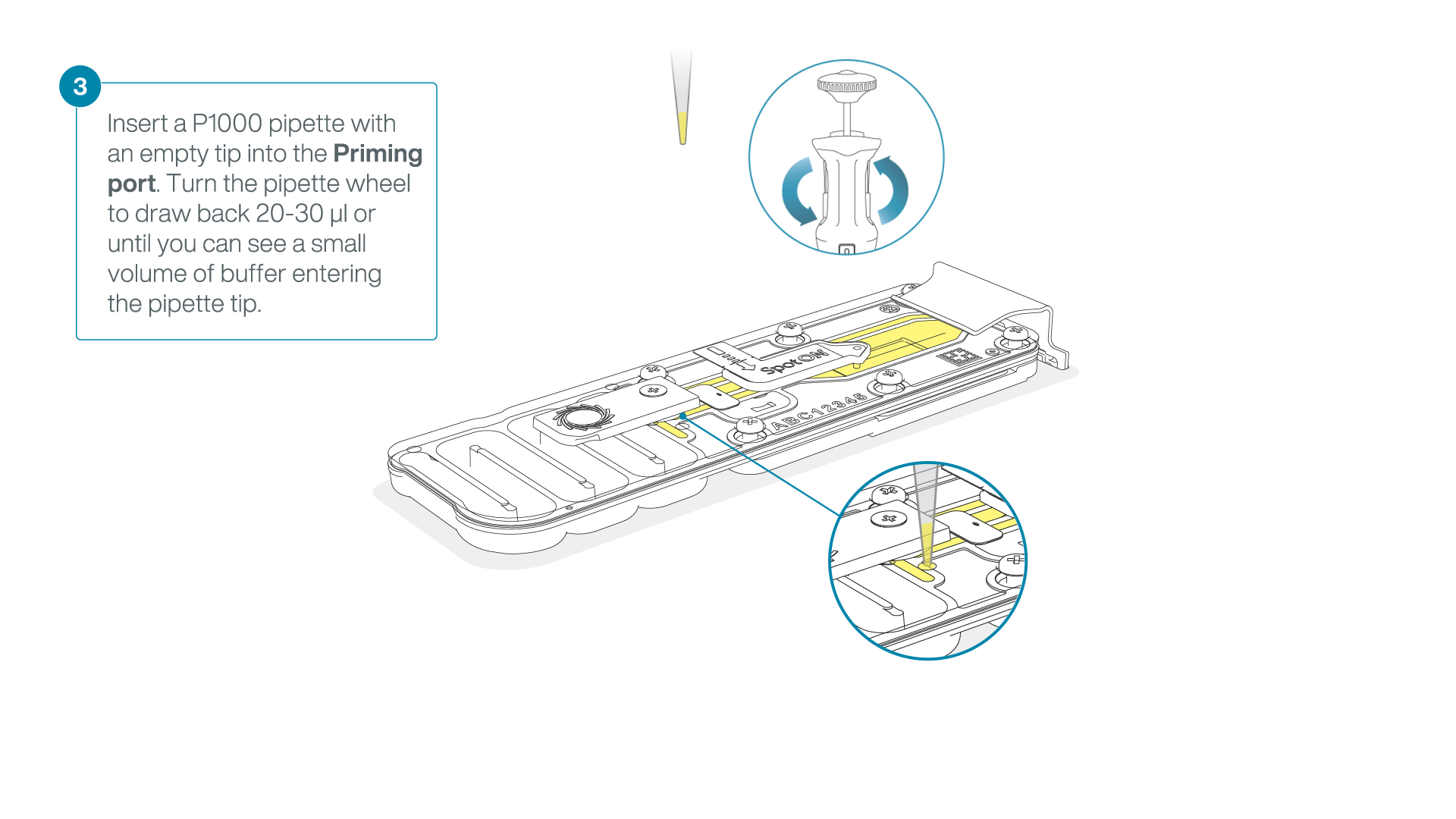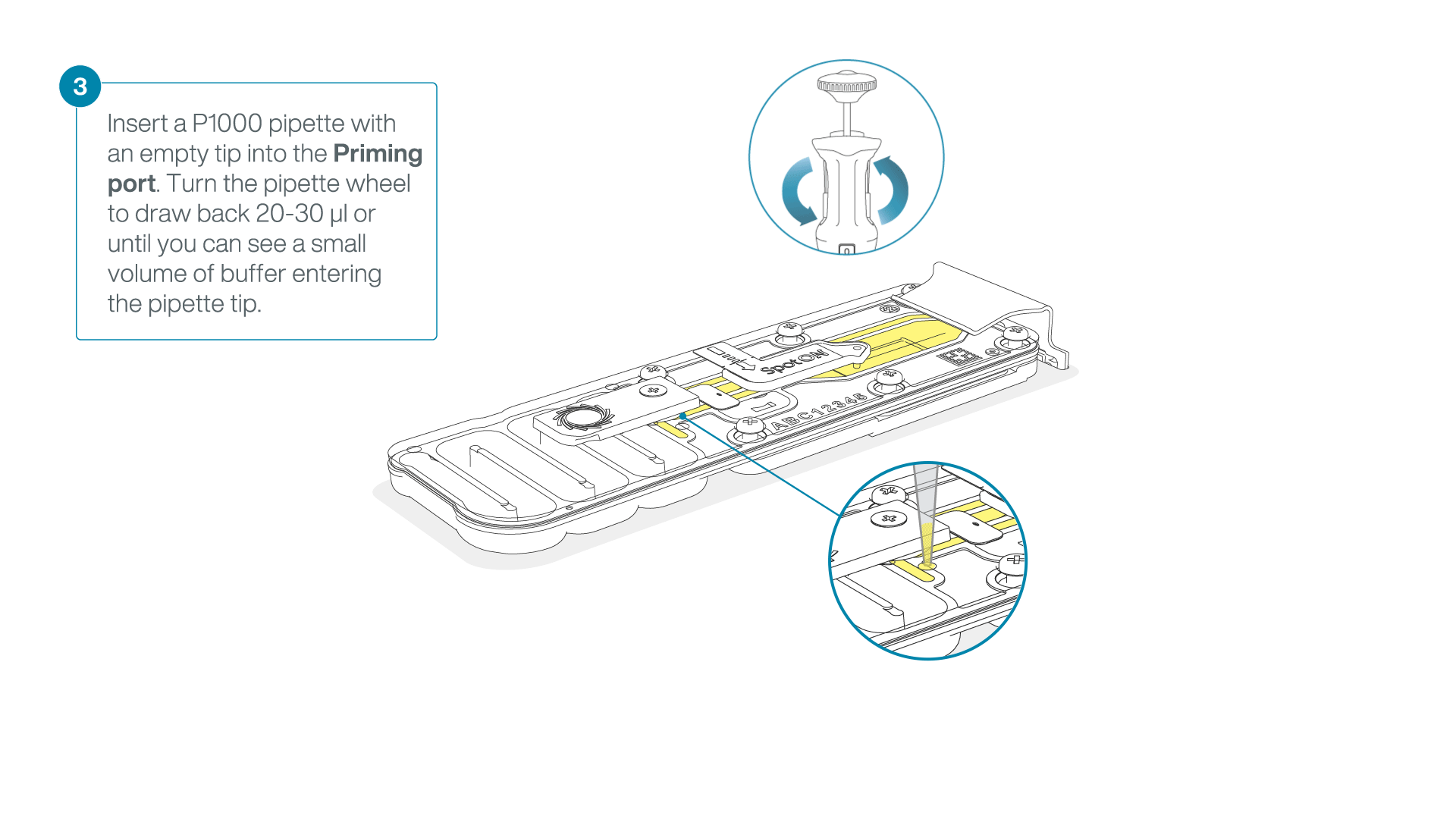
After opening the priming port, check for a small air bubble under the cover. Draw back a small volume to remove any bubbles:
- Set a P1000 pipette to 200 µl
- Insert the tip into the priming port
- Turn the wheel until the dial shows 220-230 µl, to draw back 20-30 µl, or until you can see a small volume of buffer entering the pipette tip
Note: Visually check that there is continuous buffer from the priming port across the sensor array.
-
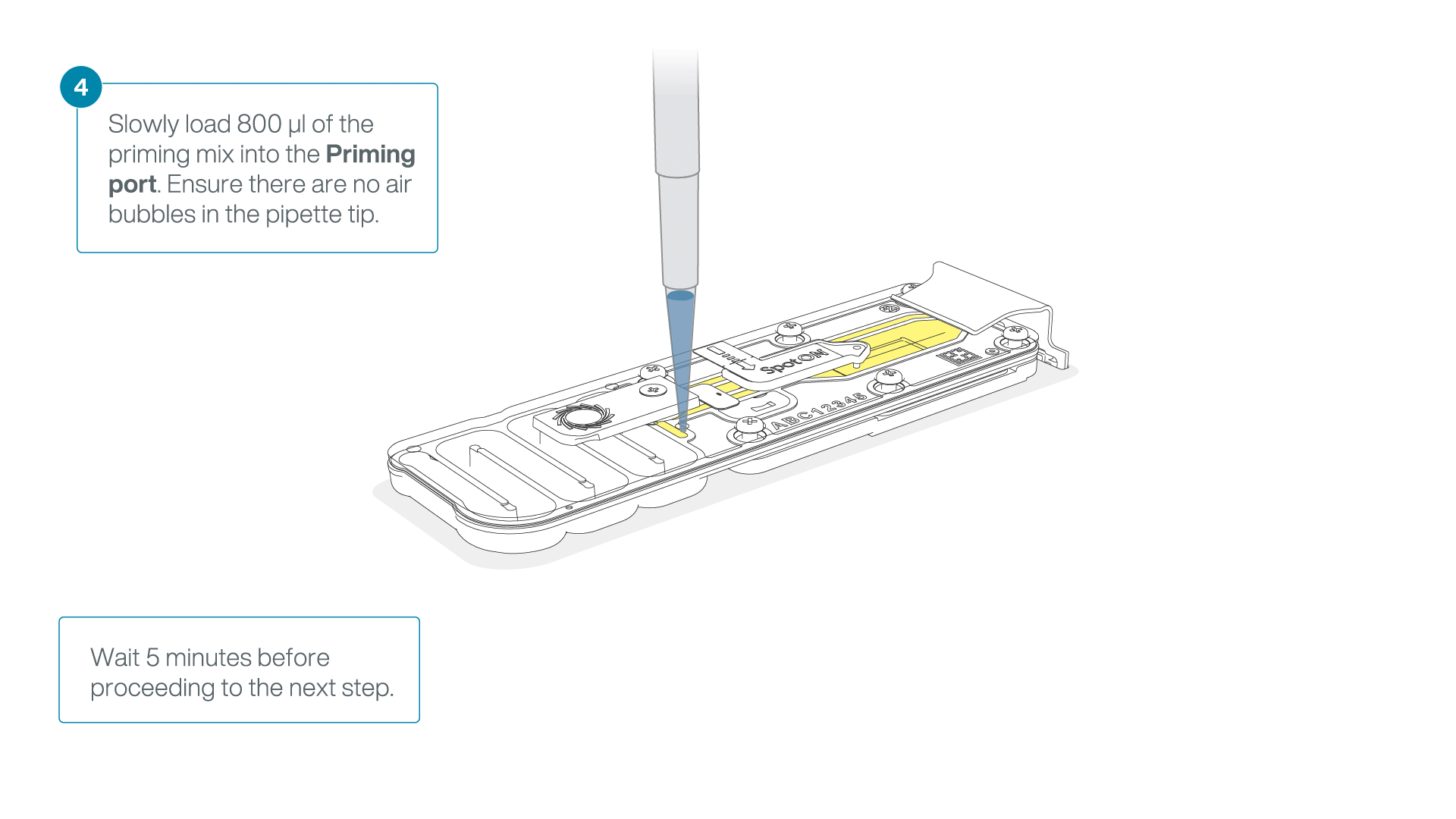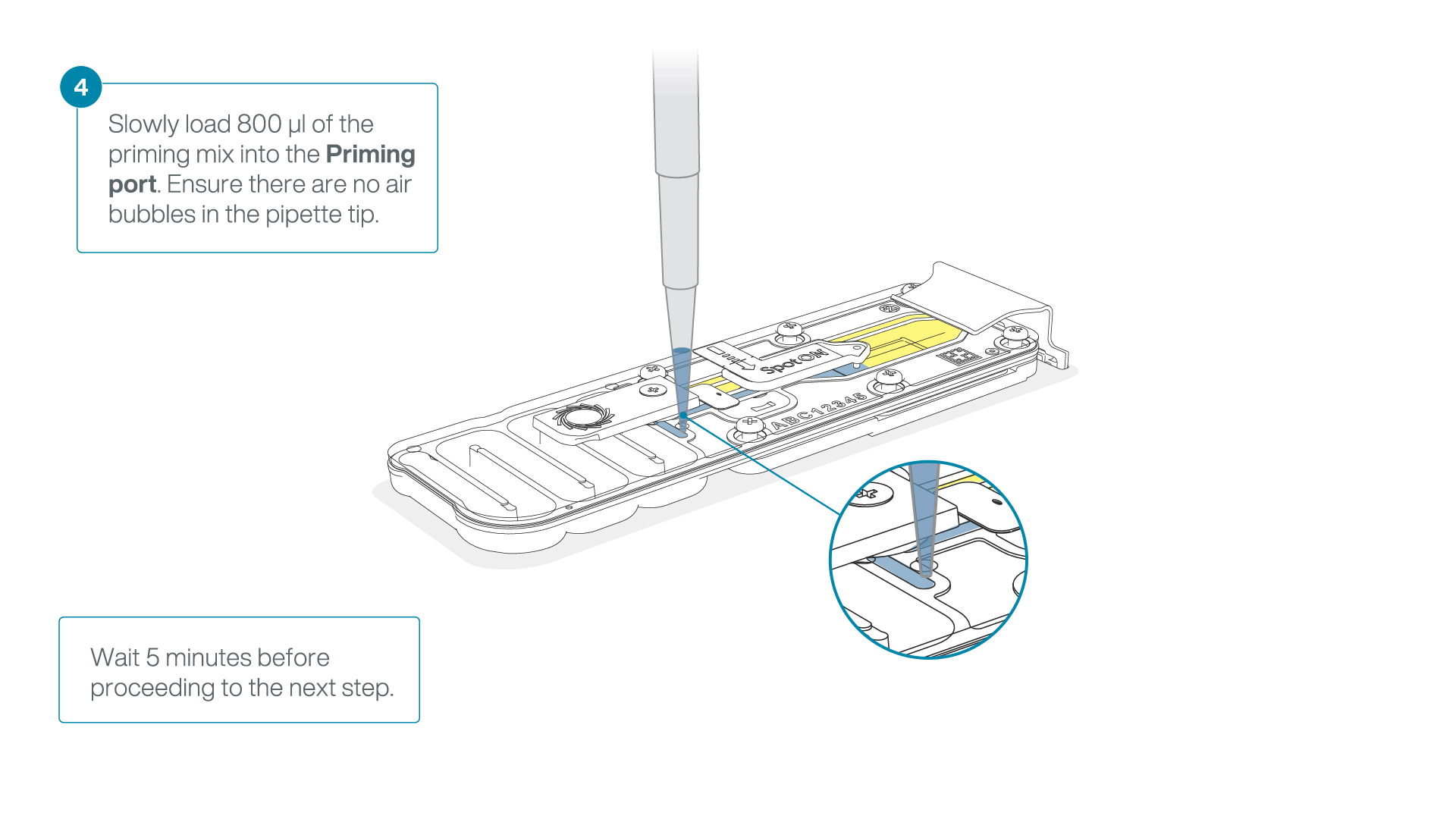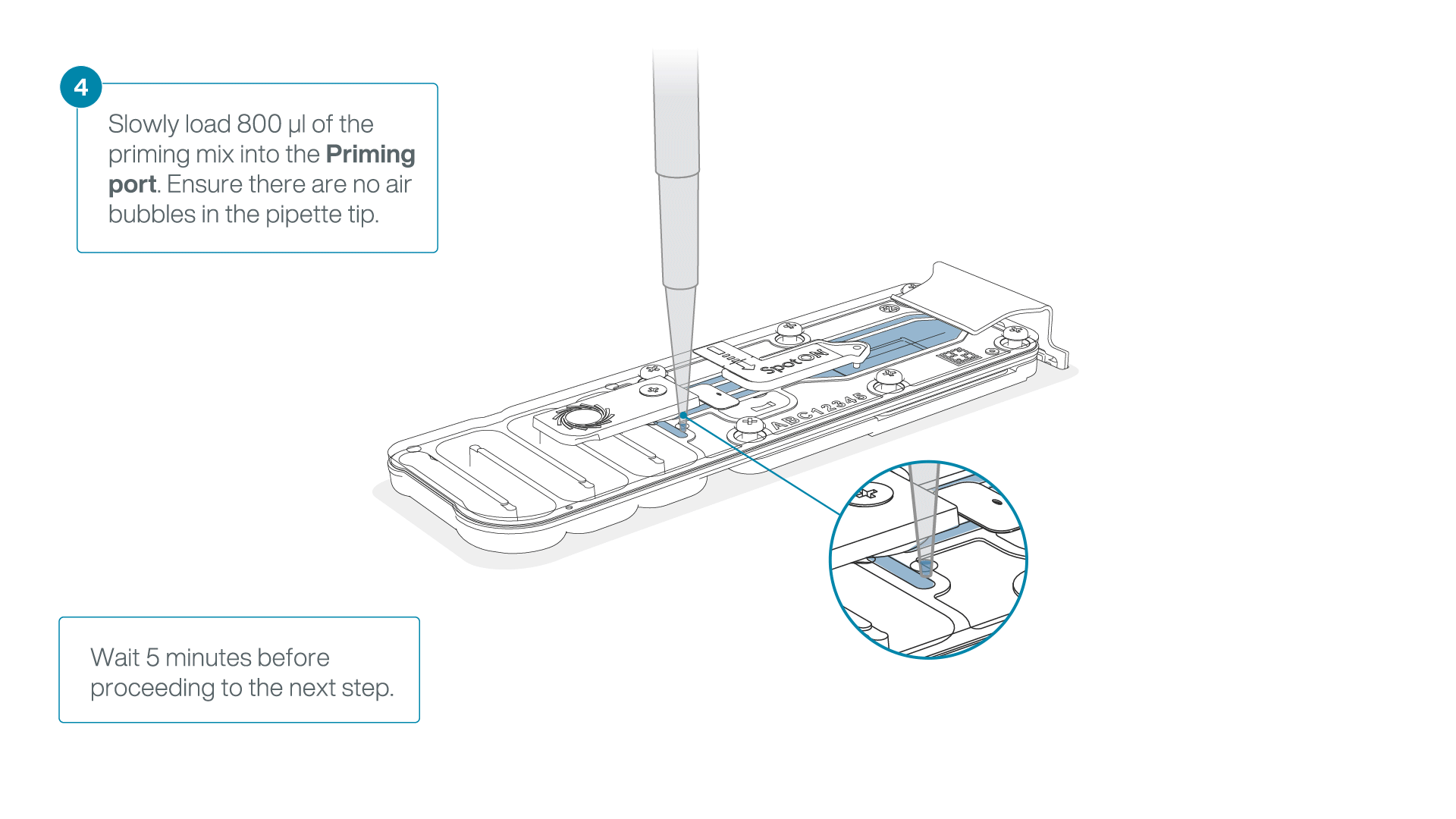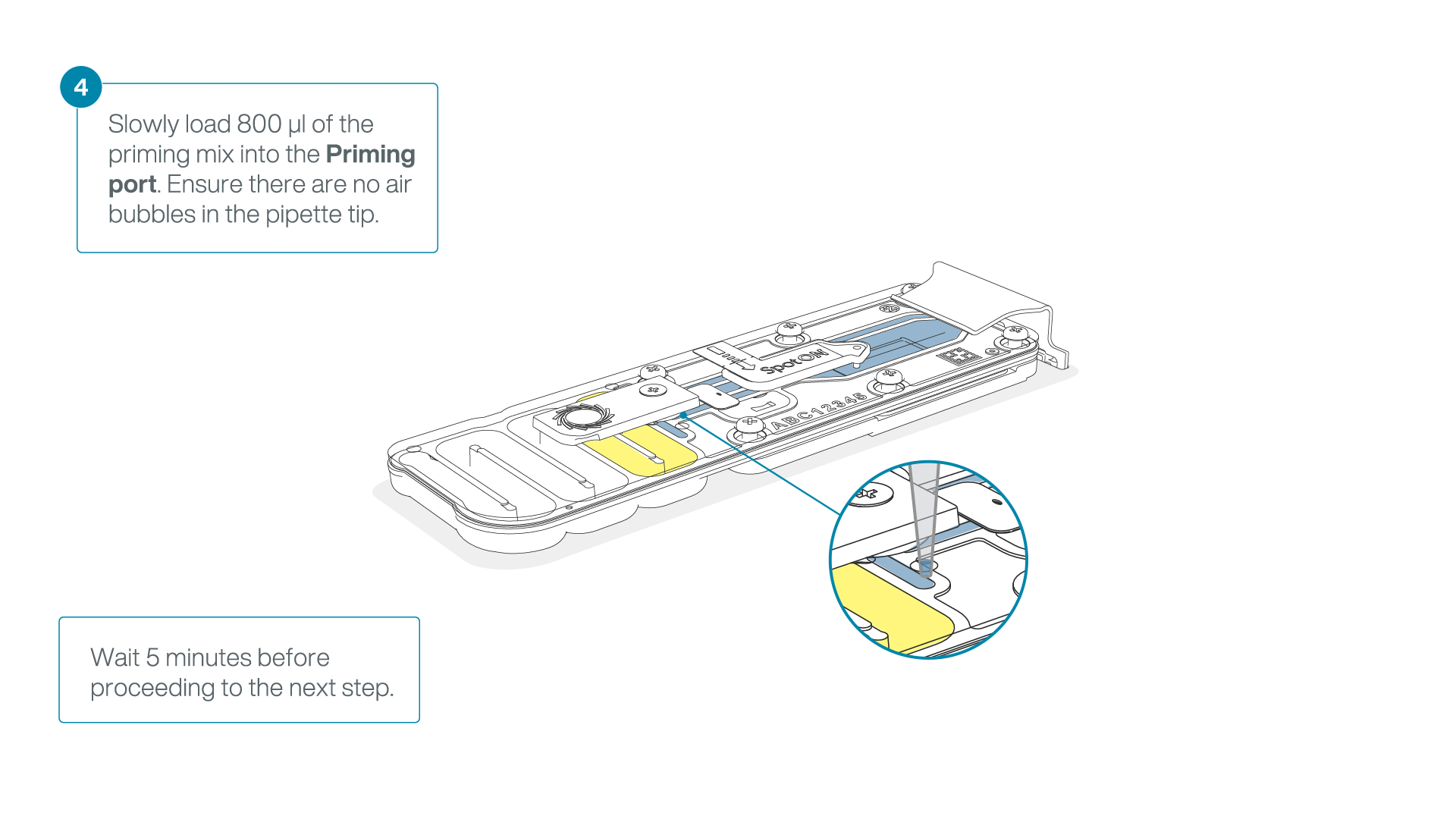
Load 800 µl of the priming mix into the flow cell via the priming port, avoiding the introduction of air bubbles. Wait for five minutes. During this time, prepare the library for loading by following the steps below.
-
Thoroughly mix the contents of the Loading Beads (LB) by pipetting.
-
In a new tube, prepare the library for loading as follows:
Reagent Volume per flow cell Sequencing Buffer (SQB) 37.5 µl Loading Beads (LB), mixed immediately before use 25.5 µl DNA library 12 µl Total 75 µl Note: Load the library onto the flow cell immediately after adding the Sequencing Buffer (SQB) and Loading Beads (LB) because the fuel in the buffer will start to be consumed by the adapter.
-
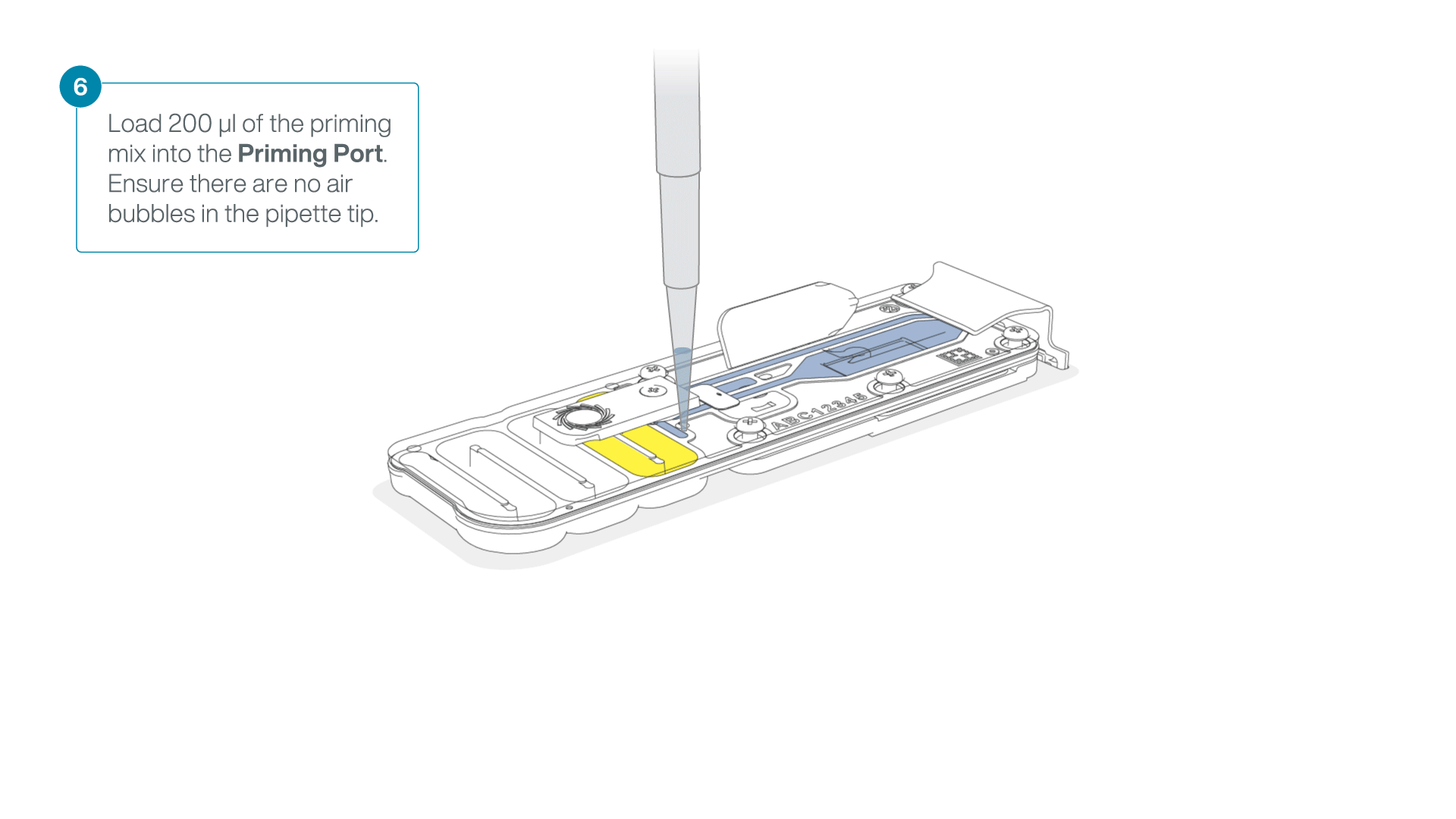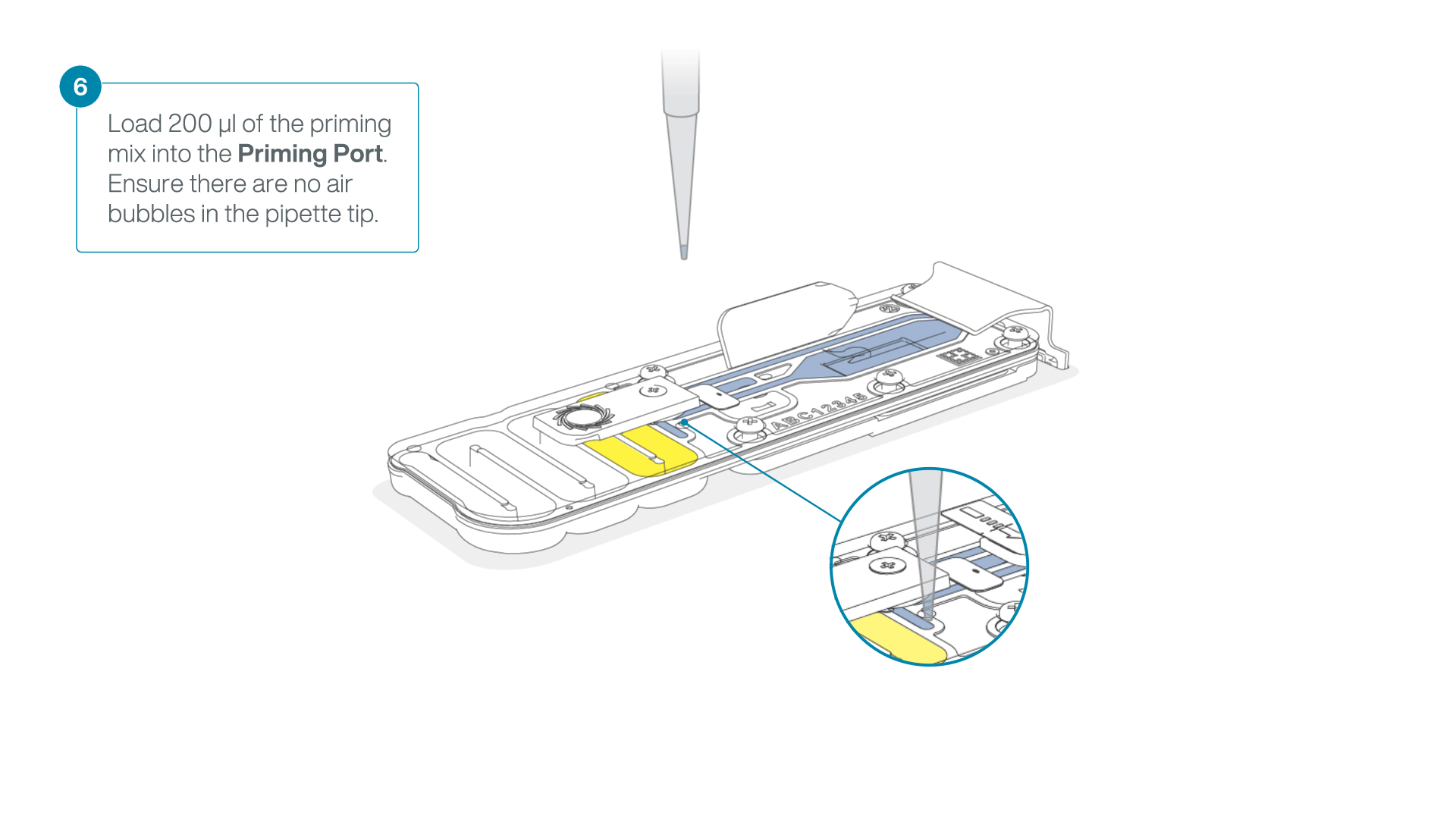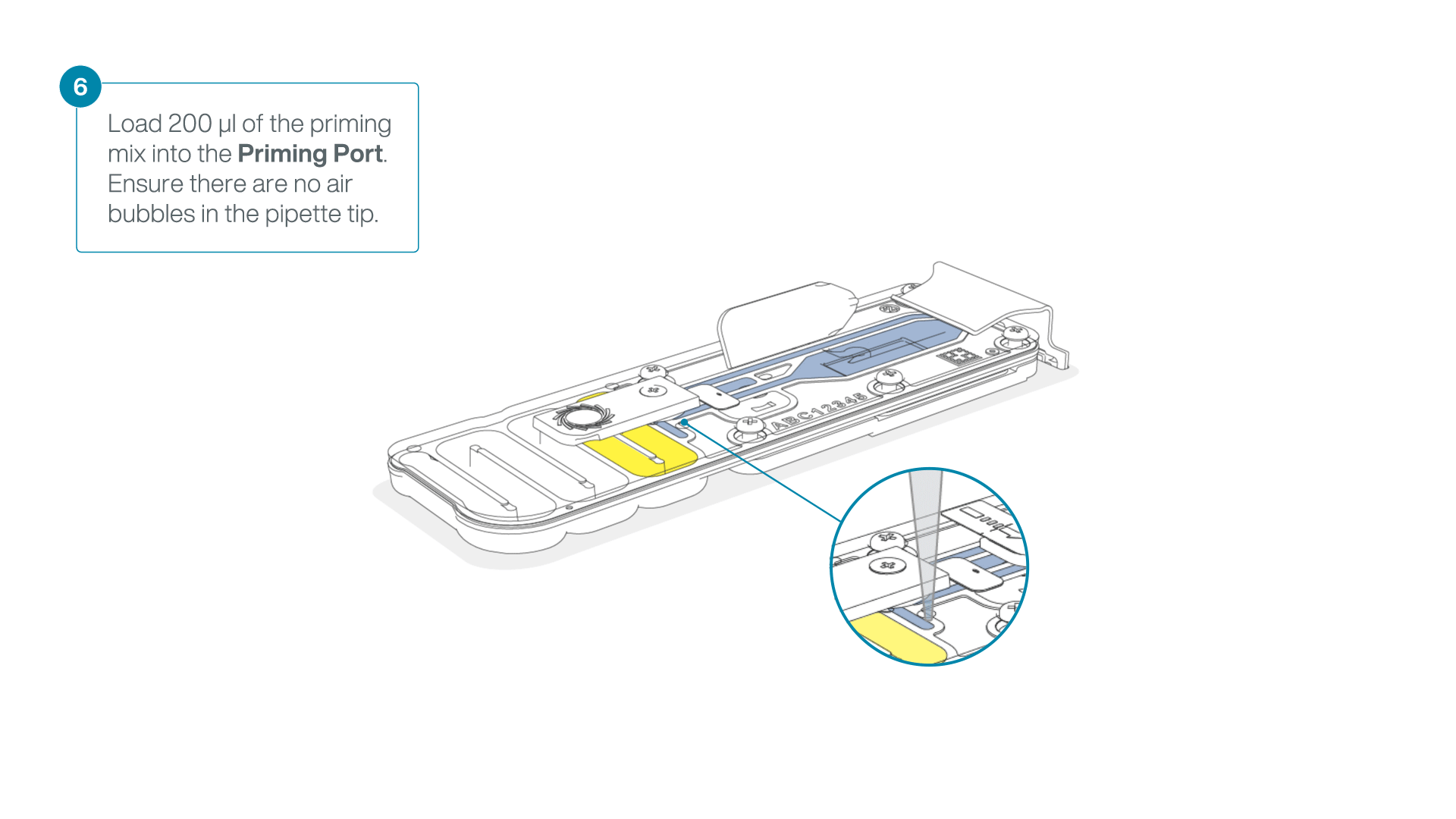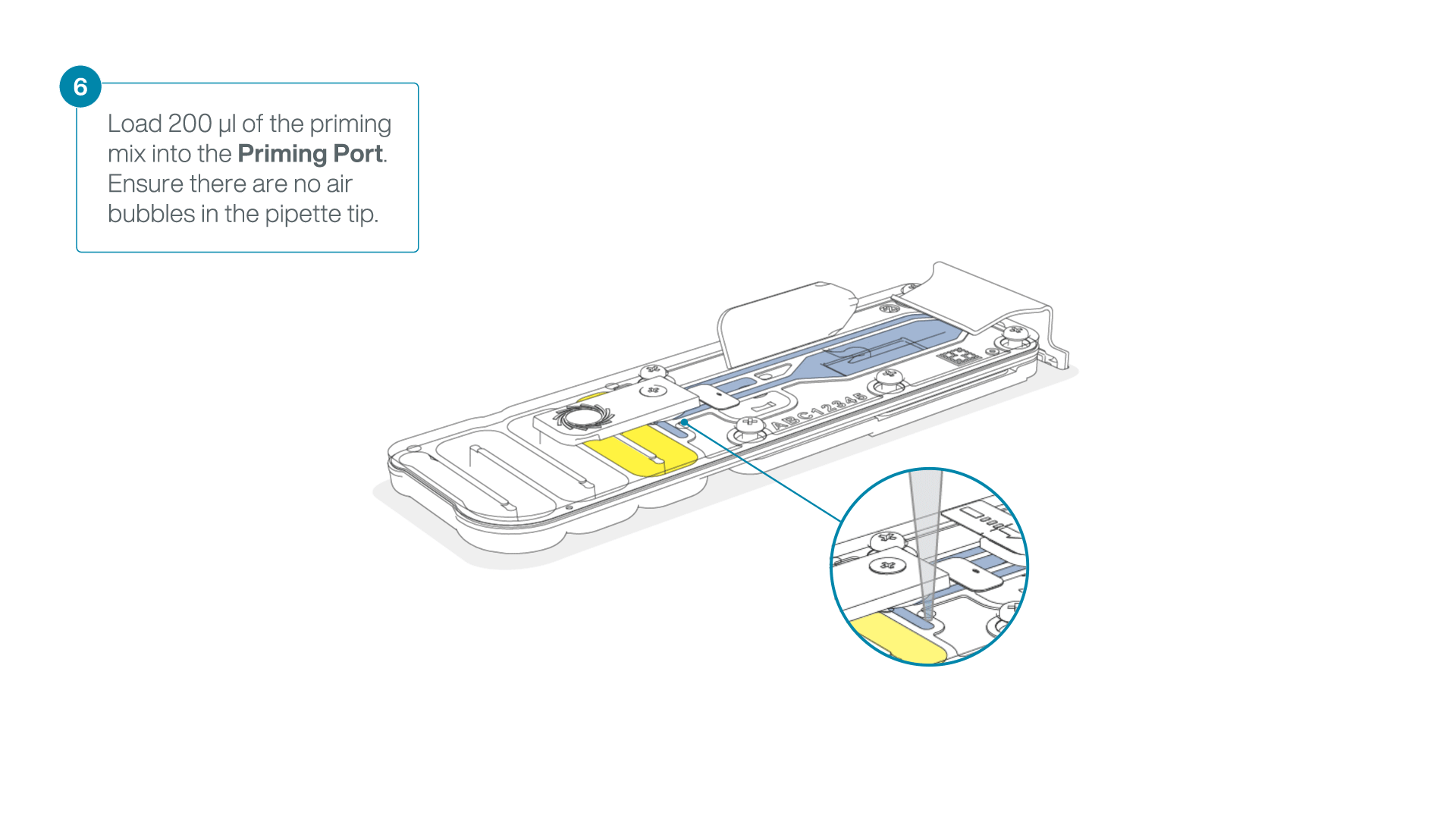
Complete the flow cell priming:
- Gently lift the SpotON sample port cover to make the SpotON sample port accessible.
- Load 200 µl of the priming mix into the flow cell priming port (not the SpotON sample port), avoiding the introduction of air bubbles.

-
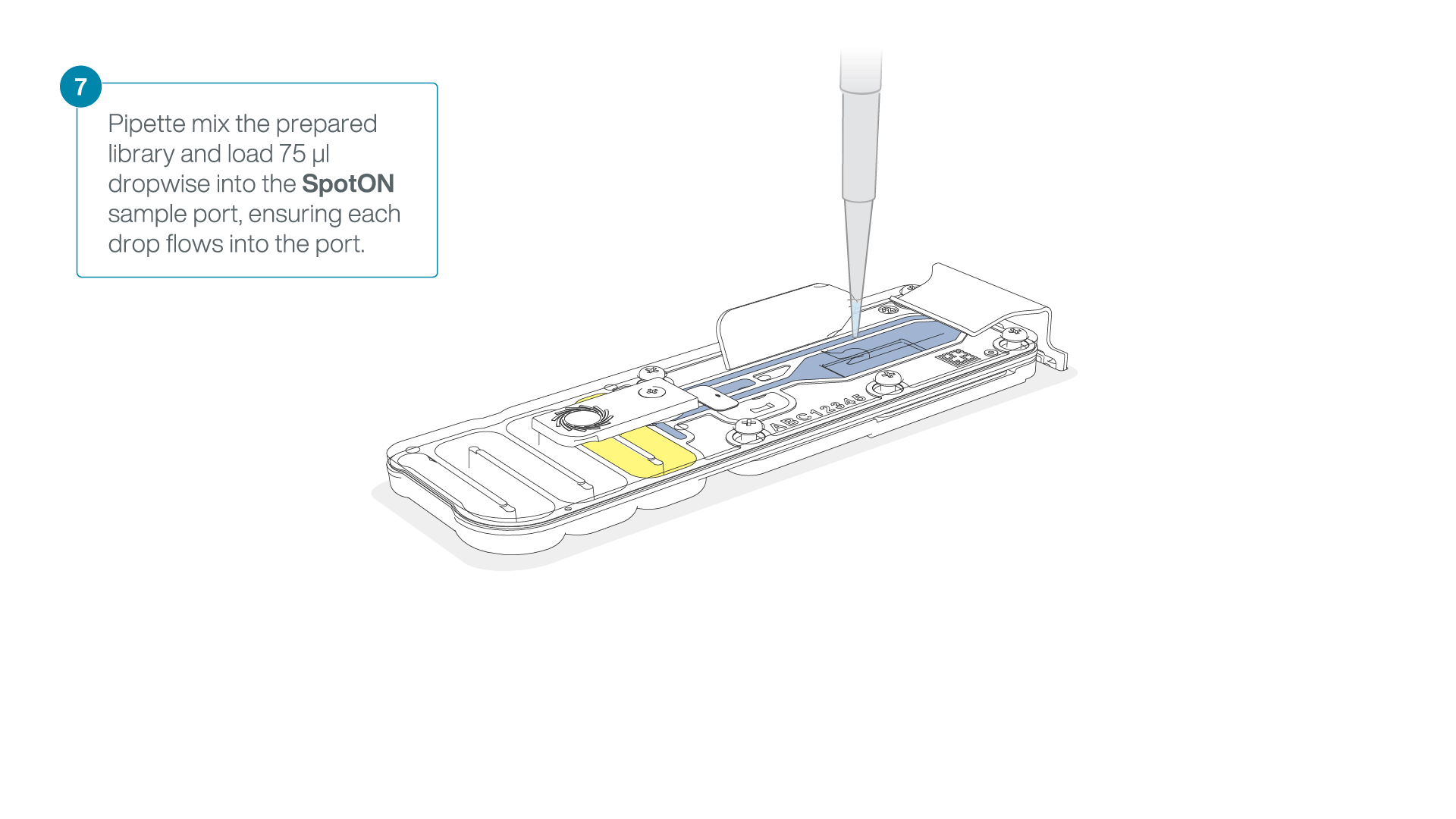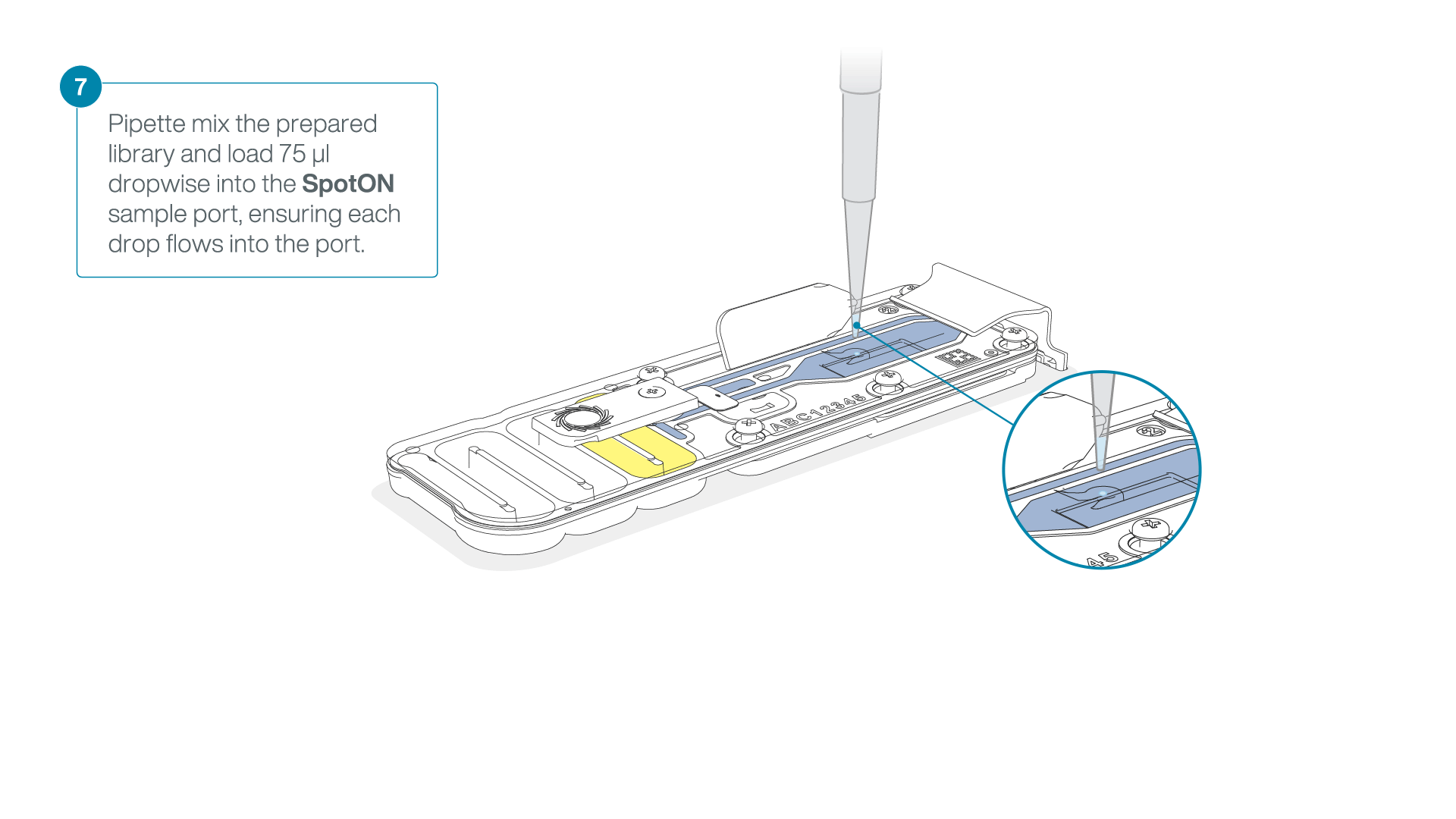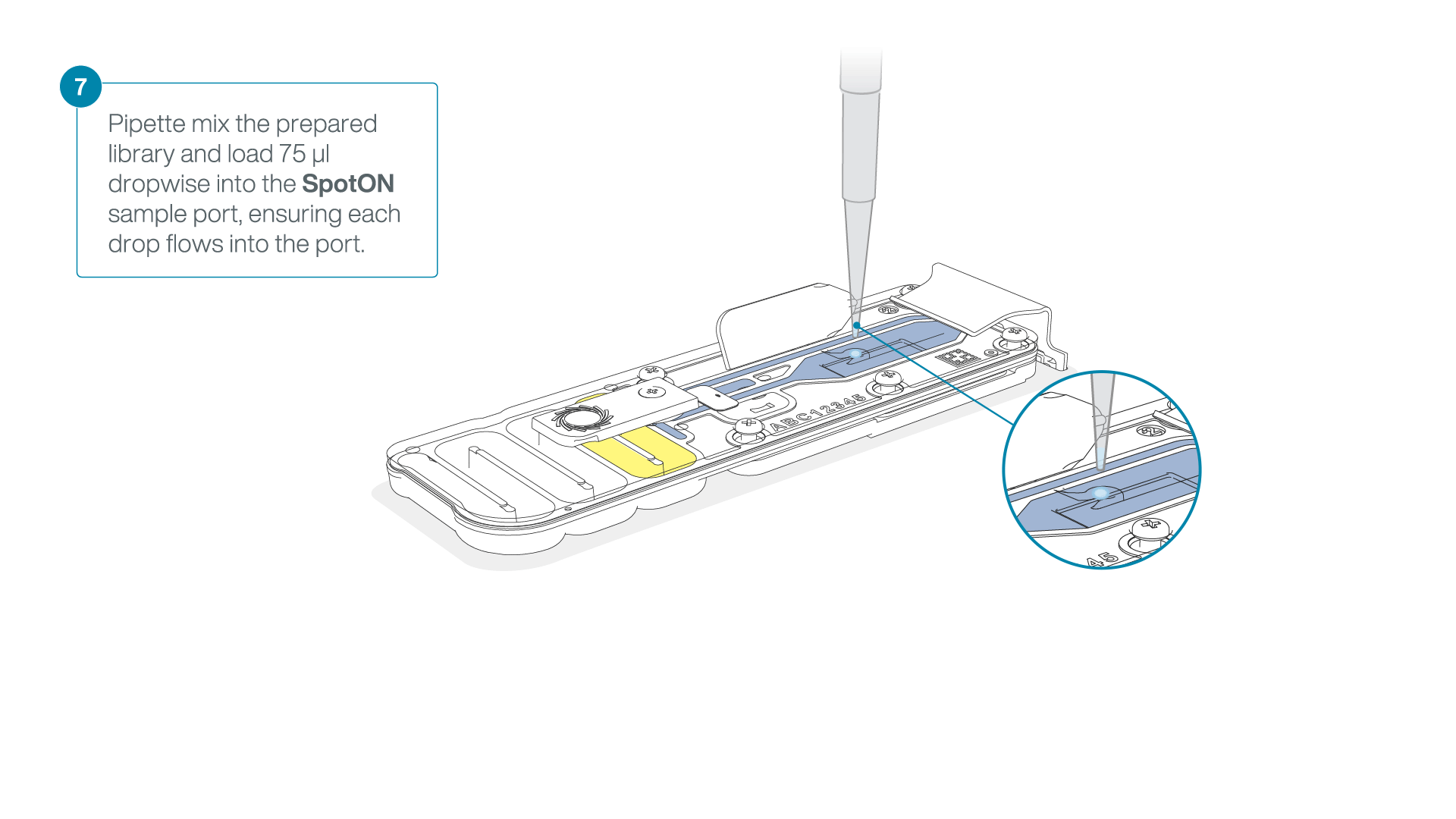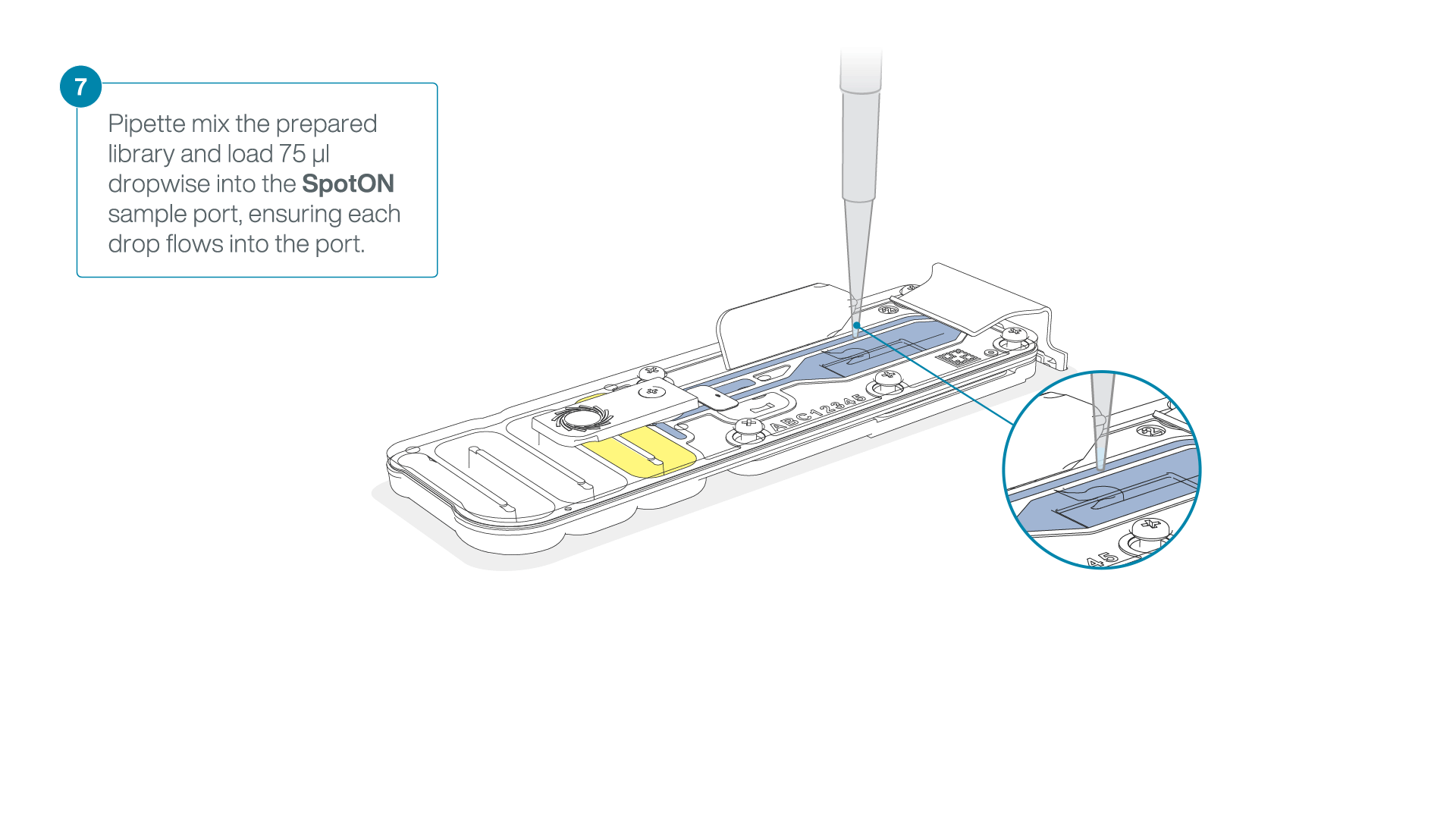
Mix the prepared library gently by pipetting up and down just prior to loading.
-
Add 75 μl of the prepared library to the flow cell via the SpotON sample port in a dropwise fashion. Ensure each drop flows into the port before adding the next.
-
Gently replace the SpotON sample port cover, making sure the bung enters the SpotON port, close the priming port and replace the MinION device lid.












Sequencing and data analysis
Data acquisition and basecalling
Data acquisition and basecalling
-
Overview of nanopore data analysis
For a full overview of nanopore data analysis, which includes options for basecalling and post-basecalling analysis, please refer to the Data Analysis document.
-
How to start sequencing
The sequencing device control, data acquisition and real-time basecalling are carried out by the MinKNOW software. Please ensure MinKNOW is installed on your computer or device. There are multiple options for how to carry out sequencing:
1. Data acquisition and basecalling in real-time using MinKNOW on a computer
Follow the instructions in the MinKNOW protocol beginning from the "Starting a sequencing run" section until the end of the "Completing a MinKNOW run" section.
2. Data acquisition and basecalling in real-time using the MinION Mk1B/Mk1D device
Follow the instructions in the MinION Mk1B user manual or the MinION Mk1D user manual.
3. Data acquisition and basecalling in real-time using the MinION Mk1C device
Follow the instructions in the MinION Mk1C user manual.
4. Data acquisition and basecalling in real-time using the GridION device
Follow the instructions in the GridION user manual.
5. Data acquisition and basecalling in real-time using the PromethION device
Follow the instructions in the PromethION user manual or the PromethION 2 Solo user manual.
6. Data acquisition using MinKNOW on a computer and basecalling at a later time using MinKNOW
Follow the instructions in the MinKNOW protocol beginning from the "Starting a sequencing run" section until the end of the "Completing a MinKNOW run" section. When setting your experiment parameters, set the Basecalling tab to OFF. After the sequencing experiment has completed, follow the instructions in the Post-run analysis section of the MinKNOW protocol.
-
When setting the sequencing parameters in MinKNOW, in the Basecalling set barcoding as Enabled, and in the barcoding options, toggle Barcode both ends, Mid-read barcodes and Override minimum mid barcoding score to ON and set Minimum mid barcoding score to 50.
Optional: basecalling and/or demultiplexing of sequences can be performed using the stand-alone Guppy software.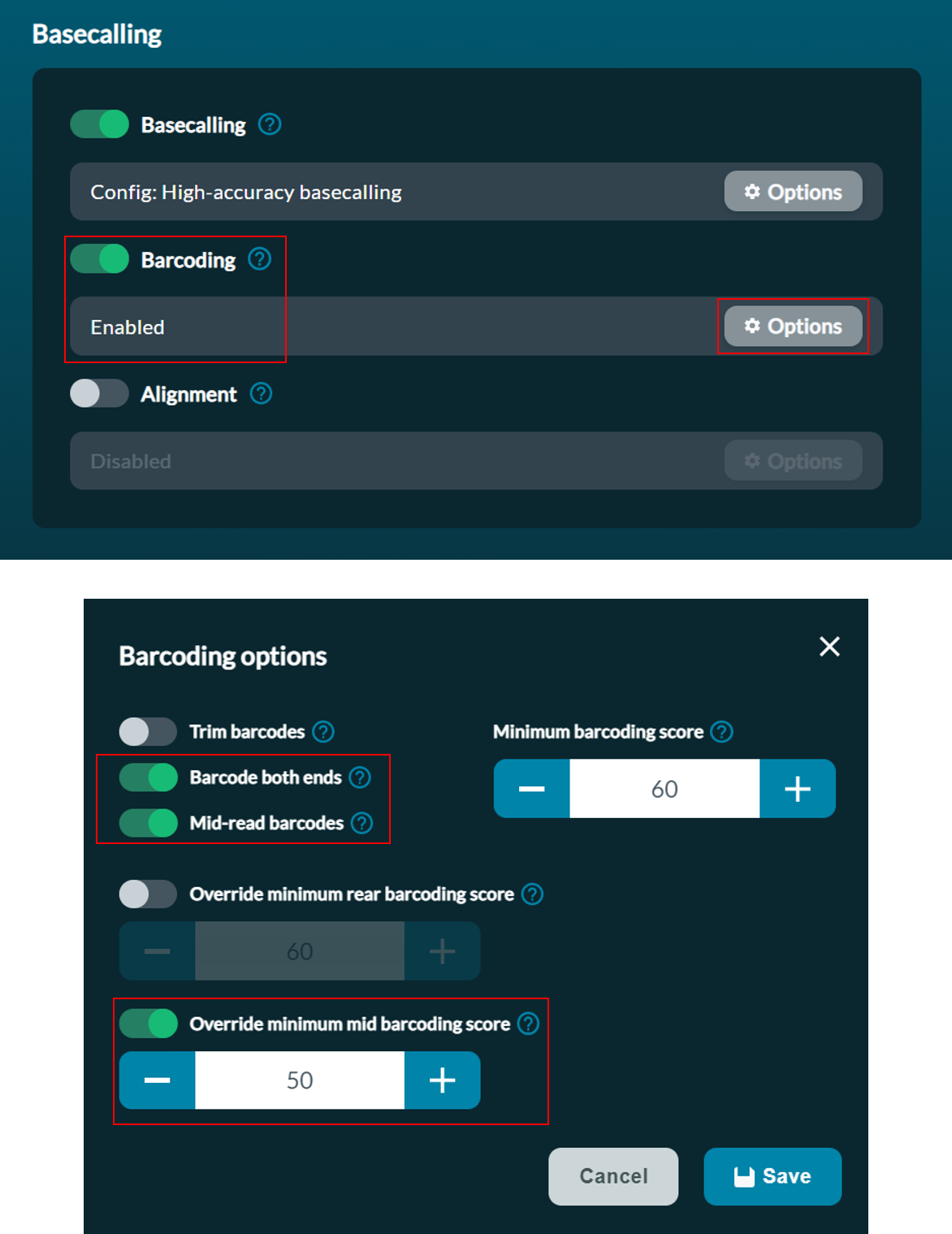

Downstream analysis and expected results
Downstream analysis and expected results
-
Recommended analysis pipeline
The recommended workflows for the bioinformatics analyses are provided by the ARTIC network and are documented on their web pages at https://artic.network/ncov-2019/ncov2019-bioinformatics-sop.html.
The reference guided genome assembly and variant calling are also performed according to the bioinformatics protocol provided by the ARTIC network. Their best practices guide uses the software contained within the FieldBioinformatics project on GitHub.
This workflow uses only the basecalled FASTQ files to perform a high-quality reference-guided assembly of the SARS-CoV-2 genome. Sequenced reads are re-demultiplexed with the requirement that reads must contain a barcode at both ends of the sequence (this only applies to the Classic and Eco PCR tiling of SARS-CoV-2 protocols but not the Rapid Barcoding PCR tiling of SARS-CoV-2), and must not contain internal barcodes. The reads are mapped to the reference genome, primer sequences are excluded and the consensus sequence is polished. The Medaka software is used to call single-nucleotide variants while the ARTIC software reports the high-quality consensus sequence from the workflow.
-
Expected results
Here, results are shown based on human coronavirus 229E spiked into 100 ng of human RNA derived from GM12878 cell line. 10 pg–0.001 pg of viral RNA obtained from ATCC was spiked into the human RNA and human-only and reverse transcription negative controls were carried through the prep to sequencing. Every sample underwent 30 and 35 cycles of PCR to determine sensitivity and specificity guidelines, as well as the expected amplicon drop-out rate for each sample.
Note: The viral RNA from ATCC is generated from cell lines infected with human coronavirus 229E. The RNA supplied is total RNA extracted from the cell lines and includes both human and viral RNA. Therefore, the levels of sensitivity are likely to be higher than those reported here.
-
Sample balancing
The graph below shows the expected sequence balancing if the protocol is followed. Here, equal masses went into the end-prep and native barcode ligation prior to pooling by equal mass for adapter ligation.
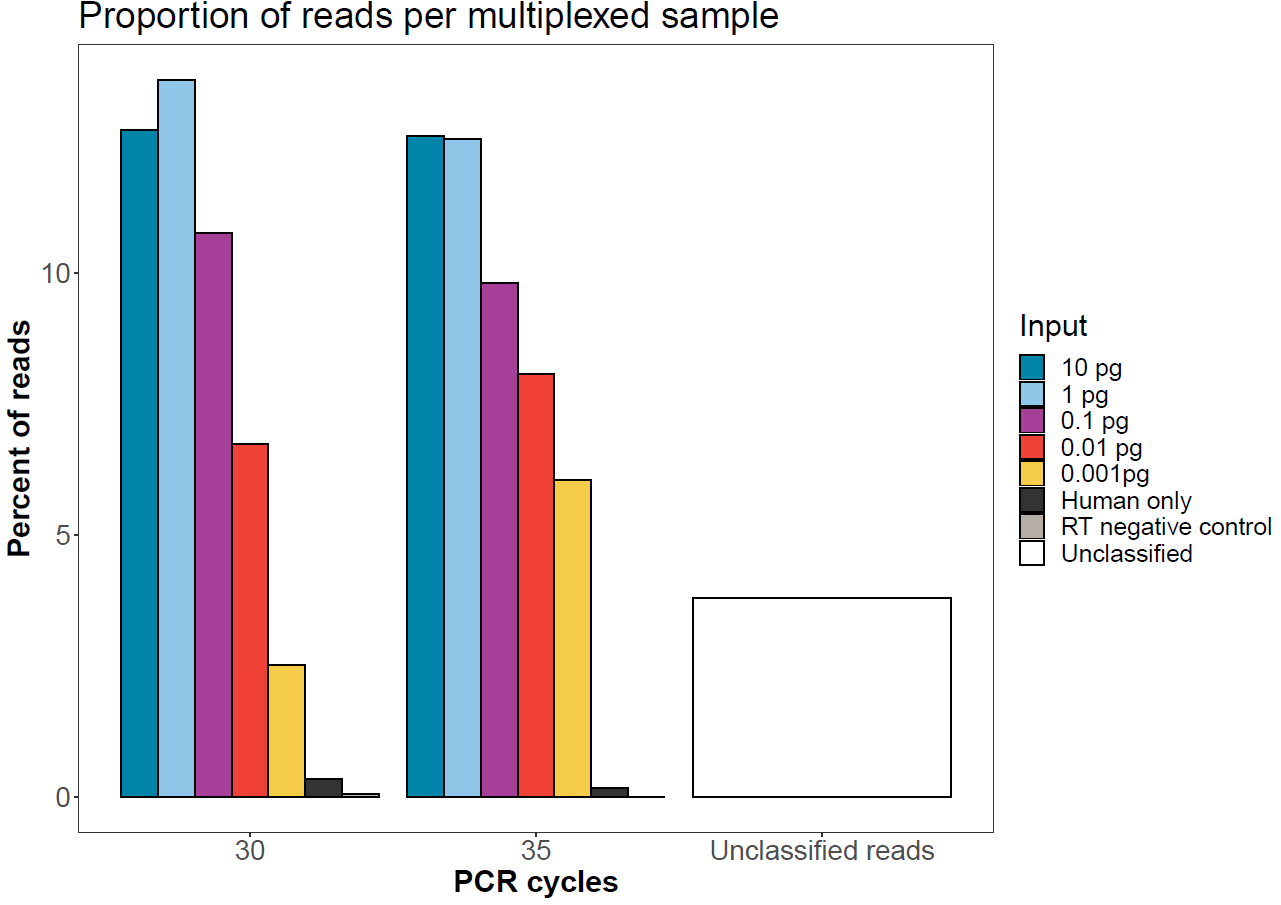
Figure 3. Number of reads per sample after native barcode demultiplexing in MinKNOW. All 14 samples were run on a single flow cell. -
On-target rate
Sequences from each demultiplexed sample were aligned to the human coronavirus 229E genome using minimap2. The proportion of primary alignments per sample are reported below.
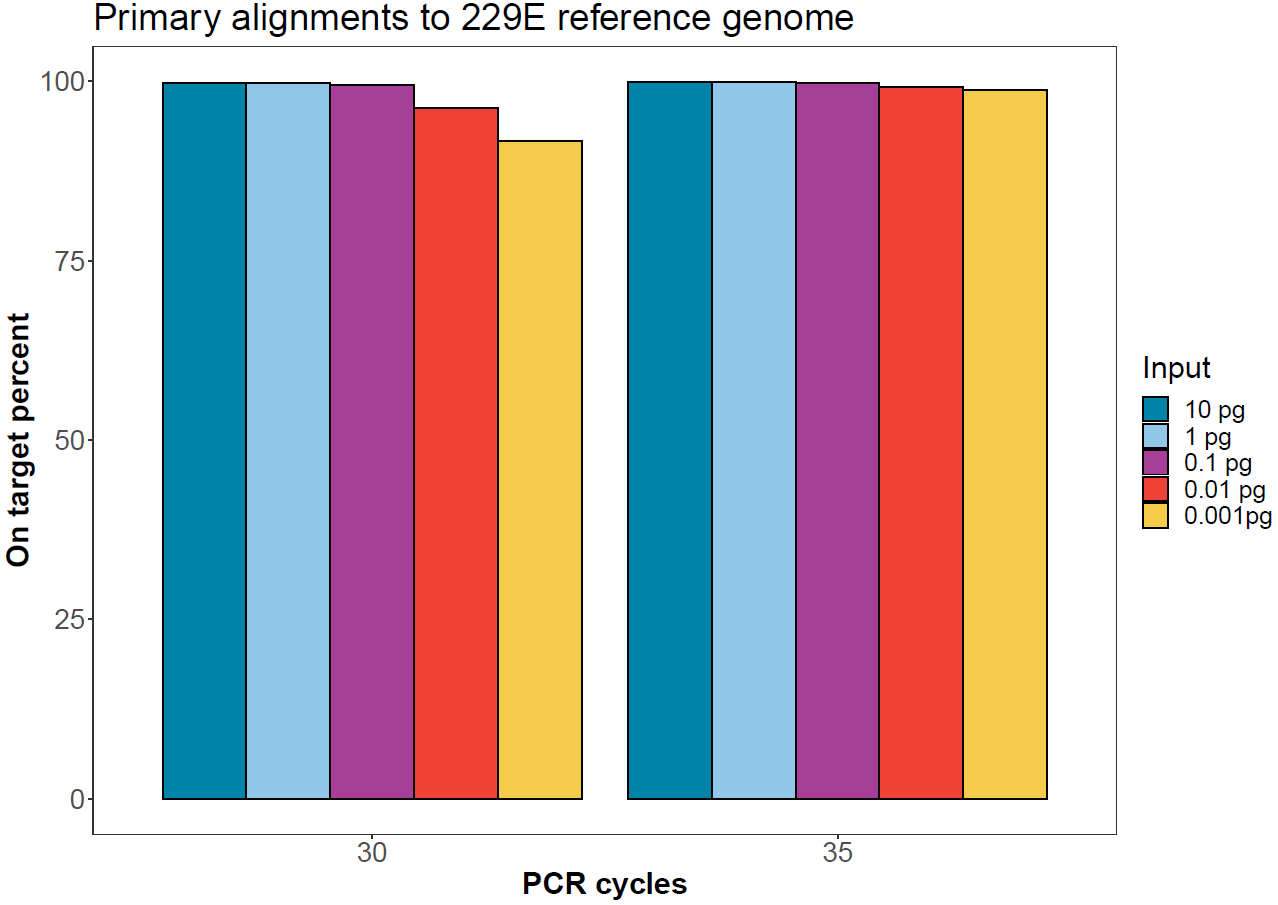
Figure 4. Proportion of reads for each sample aligning to the human coronavirus 229E reference genome. -
Assessment of negative controls
After 12 hours of sequencing, the number of reads from the negative control samples aligning to the viral reference genome is shown in the graph below and is compared with the absolute number of sequences aligning to the lowest input (0.001 pg).
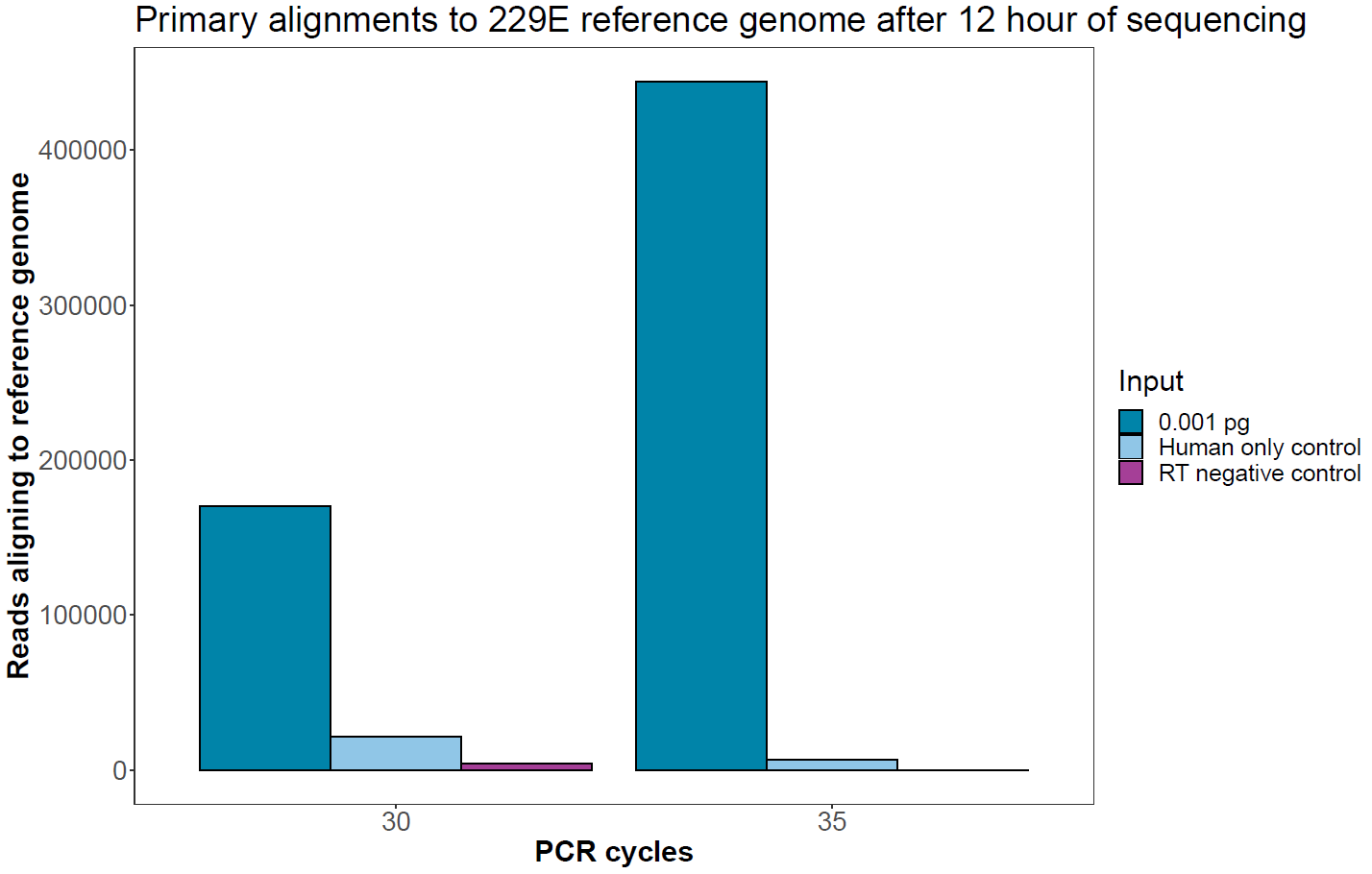
Figure 5. Absolute number of reads aligning to the human coronavirus 229E reference genome in the negative controls compared with the lowest input of viral RNA. Sequencing was carried out for 12 hours to pick up low levels of sequences assigned to barcodes representing these samples. -
Target coverage for different PCR cycles and viral load
To assess the impact of PCR dropout with lowering input viral load and increasing PCR cycles, Mosdepth was used to calculate the proportion of the viral genome covered to different depth levels. These numbers were calculated after 12 hours of sequencing with 14 samples multiplexed.
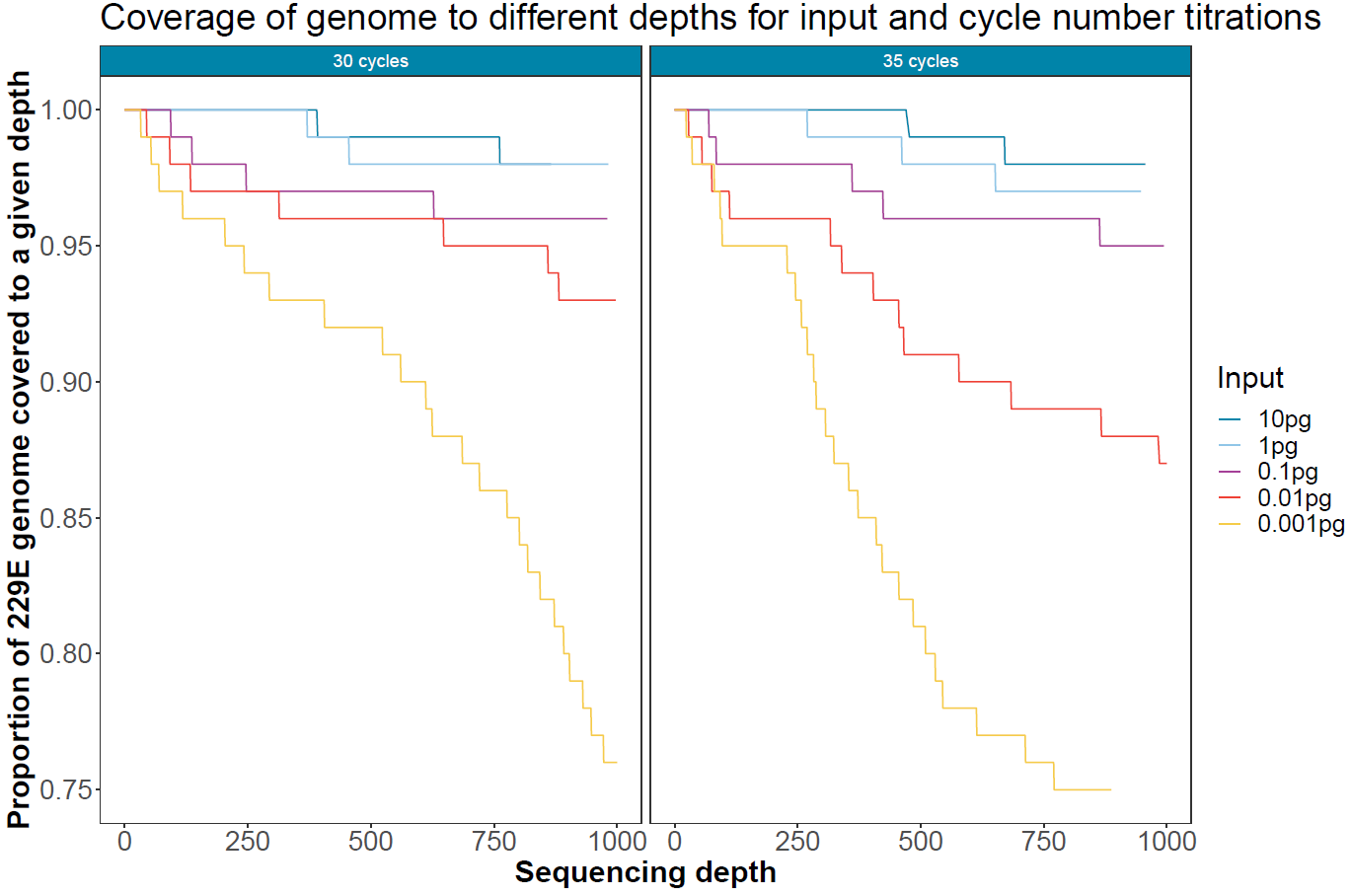
Figure 6. Coverage and depth of the human coronavirus 229E genome for different input quantities of viral RNA and different cycle numbers after 12 hours of sequencing on a single flow cell. -
How much sequencing is required?
This is unknown in real clinical samples. The graph below can be used to determine the proportion of the genome that could be covered to a given depth with different numbers of reads (30 cycles) at different input amounts in a background of 100 ng human RNA.
Note: this is absolute depth.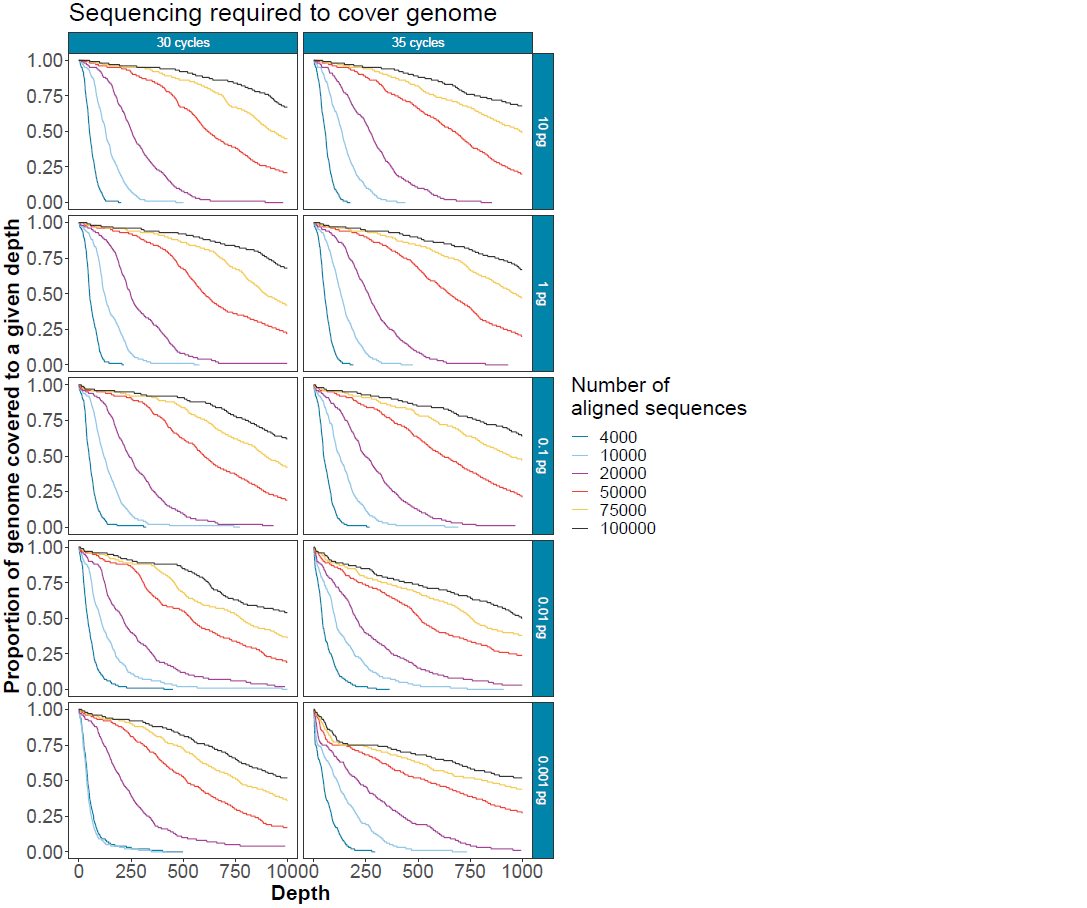
Figure 7. Subsampled sequences to give an indication of the depth of sequencing achievable covering different amounts of the human coronavirus 229E genome. Input quantities and cycle number titrations show that high cycle numbers should be avoided where possible to minimise amplicon drop out.This protocol provides amplification of low copy number viral genomes in a tiled method with low off-target amplification and minimal cross-contamination between samples. With <60 copies per reaction (0.001 pg viral input) in 100 ng background human RNA, under ideal circumstances, one should expect to cover >75% of the targeted genome at a depth of 200X within under 50,000 reads in the samples with the lowest viral titre and <20,000 reads in those with a higher viral titre.





Flow cell reuse and returns
Flow cell reuse and returns
- Materials
-
- Flow Cell Wash Kit (EXP-WSH004)
-
After your sequencing experiment is complete, if you would like to reuse the flow cell, please follow the Flow Cell Wash Kit protocol and store the washed flow cell at +2°C to +8°C.
The Flow Cell Wash Kit protocol is available on the Nanopore Community.
-
Alternatively, follow the returns procedure to send the flow cell back to Oxford Nanopore.
Instructions for returning flow cells can be found here.
Troubleshooting
Issues during DNA/RNA extraction and library preparation
Issues during DNA/RNA extraction and library preparation
-
Below is a list of the most commonly encountered issues, with some suggested causes and solutions.
We also have an FAQ section available on the Nanopore Community Support section.
If you have tried our suggested solutions and the issue still persists, please contact Technical Support via email (support@nanoporetech.com) or via LiveChat in the Nanopore Community.
-
Low sample quality
Observation Possible cause Comments and actions Low DNA purity (Nanodrop reading for DNA OD 260/280 is <1.8 and OD 260/230 is <2.0–2.2) The DNA extraction method does not provide the required purity The effects of contaminants are shown in the Contaminants document. Please try an alternative extraction method that does not result in contaminant carryover.
Consider performing an additional SPRI clean-up step.Low RNA integrity (RNA integrity number <9.5 RIN, or the rRNA band is shown as a smear on the gel) The RNA degraded during extraction Try a different RNA extraction method. For more info on RIN, please see the RNA Integrity Number document. Further information can be found in the DNA/RNA Handling page. RNA has a shorter than expected fragment length The RNA degraded during extraction Try a different RNA extraction method. For more info on RIN, please see the RNA Integrity Number document. Further information can be found in the DNA/RNA Handling page.
We recommend working in an RNase-free environment, and to keep your lab equipment RNase-free when working with RNA. -
Low DNA recovery after AMPure bead clean-up
Observation Possible cause Comments and actions Low recovery DNA loss due to a lower than intended AMPure beads-to-sample ratio 1. AMPure beads settle quickly, so ensure they are well resuspended before adding them to the sample.
2. When the AMPure beads-to-sample ratio is lower than 0.4:1, DNA fragments of any size will be lost during the clean-up.Low recovery DNA fragments are shorter than expected The lower the AMPure beads-to-sample ratio, the more stringent the selection against short fragments. Please always determine the input DNA length on an agarose gel (or other gel electrophoresis methods) and then calculate the appropriate amount of AMPure beads to use. 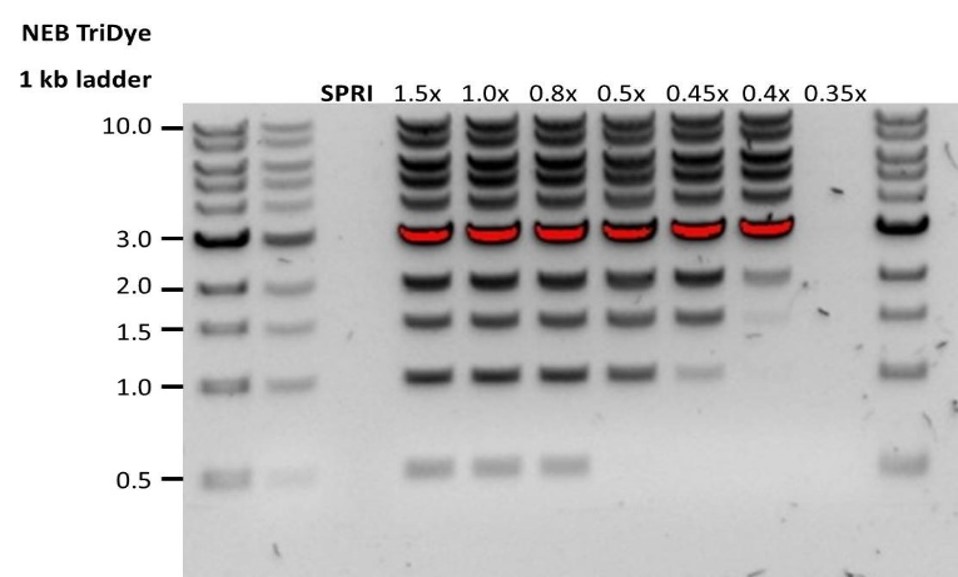
Low recovery after end-prep The wash step used ethanol <70% DNA will be eluted from the beads when using ethanol <70%. Make sure to use the correct percentage.

Issues during the sequencing run
Issues during the sequencing run
-
Below is a list of the most commonly encountered issues, with some suggested causes and solutions.
We also have an FAQ section available on the Nanopore Community Support section.
If you have tried our suggested solutions and the issue still persists, please contact Technical Support via email (support@nanoporetech.com) or via LiveChat in the Nanopore Community.
-
Fewer pores at the start of sequencing than after Flow Cell Check
Observation Possible cause Comments and actions MinKNOW reported a lower number of pores at the start of sequencing than the number reported by the Flow Cell Check An air bubble was introduced into the nanopore array After the Flow Cell Check it is essential to remove any air bubbles near the priming port before priming the flow cell. If not removed, the air bubble can travel to the nanopore array and irreversibly damage the nanopores that have been exposed to air. The best practice to prevent this from happening is demonstrated in this video. MinKNOW reported a lower number of pores at the start of sequencing than the number reported by the Flow Cell Check The flow cell is not correctly inserted into the device Stop the sequencing run, remove the flow cell from the sequencing device and insert it again, checking that the flow cell is firmly seated in the device and that it has reached the target temperature. If applicable, try a different position on the device (GridION/PromethION). MinKNOW reported a lower number of pores at the start of sequencing than the number reported by the Flow Cell Check Contaminations in the library damaged or blocked the pores The pore count during the Flow Cell Check is performed using the QC DNA molecules present in the flow cell storage buffer. At the start of sequencing, the library itself is used to estimate the number of active pores. Because of this, variability of about 10% in the number of pores is expected. A significantly lower pore count reported at the start of sequencing can be due to contaminants in the library that have damaged the membranes or blocked the pores. Alternative DNA/RNA extraction or purification methods may be needed to improve the purity of the input material. The effects of contaminants are shown in the Contaminants Know-how piece. Please try an alternative extraction method that does not result in contaminant carryover. -
MinKNOW script failed
Observation Possible cause Comments and actions MinKNOW shows "Script failed" Restart the computer and then restart MinKNOW. If the issue persists, please collect the MinKNOW log files and contact Technical Support. If you do not have another sequencing device available, we recommend storing the flow cell and the loaded library at 4°C and contact Technical Support for further storage guidance. -
Pore occupancy below 40%
Observation Possible cause Comments and actions Pore occupancy <40% Not enough library was loaded on the flow cell Ensure you load the recommended amount of good quality library in the relevant library prep protocol onto your flow cell. Please quantify the library before loading and calculate mols using tools like the Promega Biomath Calculator, choosing "dsDNA: µg to pmol" Pore occupancy close to 0 The Ligation Sequencing Kit was used, and sequencing adapters did not ligate to the DNA Make sure to use the NEBNext Quick Ligation Module (E6056) and Oxford Nanopore Technologies Ligation Buffer (LNB, provided in the sequencing kit) at the sequencing adapter ligation step, and use the correct amount of each reagent. A Lambda control library can be prepared to test the integrity of the third-party reagents. Pore occupancy close to 0 The Ligation Sequencing Kit was used, and ethanol was used instead of LFB or SFB at the wash step after sequencing adapter ligation Ethanol can denature the motor protein on the sequencing adapters. Make sure the LFB or SFB buffer was used after ligation of sequencing adapters. Pore occupancy close to 0 No tether on the flow cell Tethers are adding during flow cell priming (FLT/FCT tube). Make sure FLT/FCT was added to FB/FCF before priming. -
Shorter than expected read length
Observation Possible cause Comments and actions Shorter than expected read length Unwanted fragmentation of DNA sample Read length reflects input DNA fragment length. Input DNA can be fragmented during extraction and library prep.
1. Please review the Extraction Methods in the Nanopore Community for best practice for extraction.
2. Visualise the input DNA fragment length distribution on an agarose gel before proceeding to the library prep.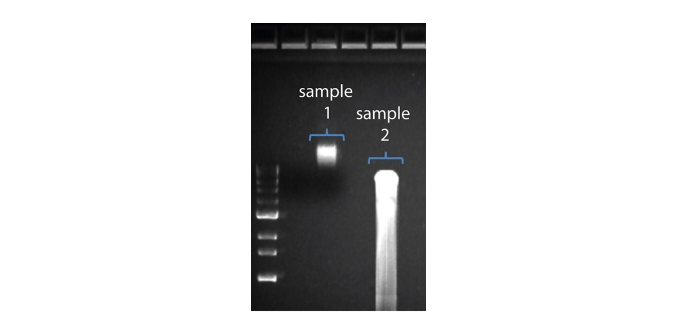 In the image above, Sample 1 is of high molecular weight, whereas Sample 2 has been fragmented.
In the image above, Sample 1 is of high molecular weight, whereas Sample 2 has been fragmented.
3. During library prep, avoid pipetting and vortexing when mixing reagents. Flicking or inverting the tube is sufficient. -
Large proportion of unavailable pores
Observation Possible cause Comments and actions Large proportion of unavailable pores (shown as blue in the channels panel and pore activity plot) 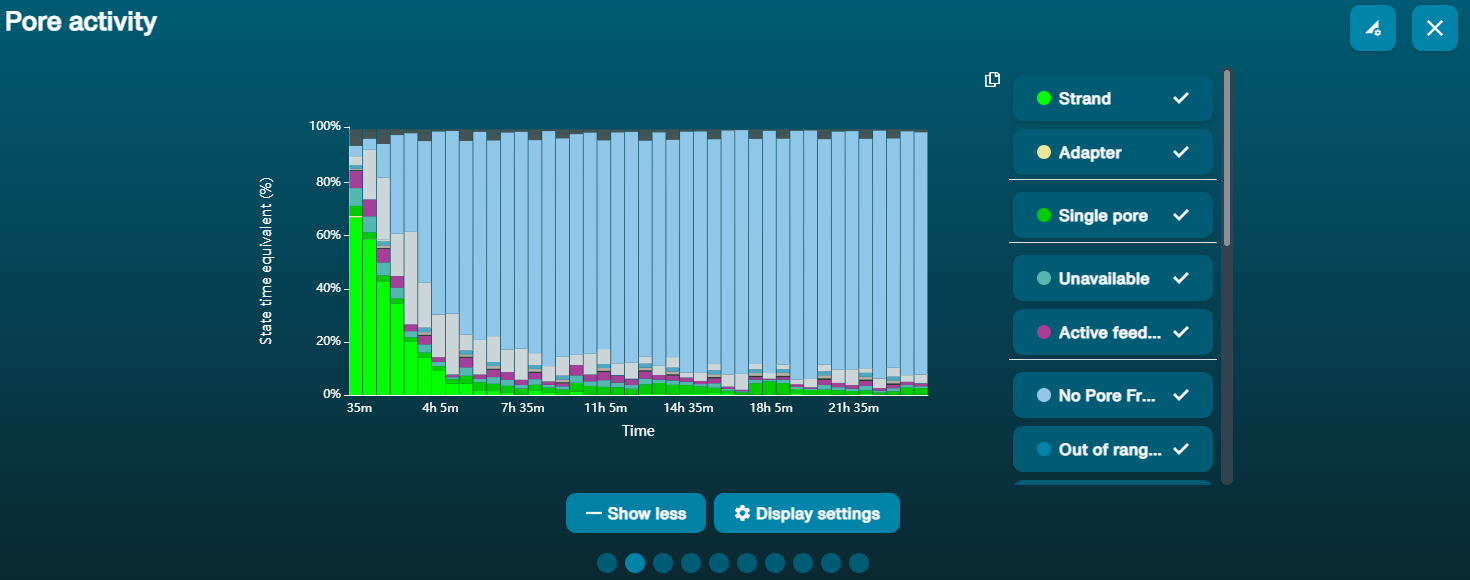 The pore activity plot above shows an increasing proportion of "unavailable" pores over time.
The pore activity plot above shows an increasing proportion of "unavailable" pores over time.Contaminants are present in the sample Some contaminants can be cleared from the pores by the unblocking function built into MinKNOW. If this is successful, the pore status will change to "sequencing pore". If the portion of unavailable pores stays large or increases:
1. A nuclease flush using the Flow Cell Wash Kit (EXP-WSH004) can be performed, or
2. Run several cycles of PCR to try and dilute any contaminants that may be causing problems. -
Large proportion of inactive pores
Observation Possible cause Comments and actions Large proportion of inactive/unavailable pores (shown as light blue in the channels panel and pore activity plot. Pores or membranes are irreversibly damaged) Air bubbles have been introduced into the flow cell Air bubbles introduced through flow cell priming and library loading can irreversibly damage the pores. Watch the Priming and loading your flow cell video for best practice Large proportion of inactive/unavailable pores Certain compounds co-purified with DNA Known compounds, include polysaccharides, typically associate with plant genomic DNA.
1. Please refer to the Plant leaf DNA extraction method.
2. Clean-up using the QIAGEN PowerClean Pro kit.
3. Perform a whole genome amplification with the original gDNA sample using the QIAGEN REPLI-g kit.Large proportion of inactive/unavailable pores Contaminants are present in the sample The effects of contaminants are shown in the Contaminants Know-how piece. Please try an alternative extraction method that does not result in contaminant carryover. -
Reduction in sequencing speed and q-score later into the run
Observation Possible cause Comments and actions Reduction in sequencing speed and q-score later into the run For Kit 9 chemistry (e.g. SQK-LSK109), fast fuel consumption is typically seen when the flow cell is overloaded with library (please see the appropriate protocol for your DNA library to see the recommendation). Add more fuel to the flow cell by following the instructions in the MinKNOW protocol. In future experiments, load lower amounts of library to the flow cell. -
Temperature fluctuation
Observation Possible cause Comments and actions Temperature fluctuation The flow cell has lost contact with the device Check that there is a heat pad covering the metal plate on the back of the flow cell. Re-insert the flow cell and press it down to make sure the connector pins are firmly in contact with the device. If the problem persists, please contact Technical Services. -
Failed to reach target temperature
Observation Possible cause Comments and actions MinKNOW shows "Failed to reach target temperature" The instrument was placed in a location that is colder than normal room temperature, or a location with poor ventilation (which leads to the flow cells overheating) MinKNOW has a default timeframe for the flow cell to reach the target temperature. Once the timeframe is exceeded, an error message will appear and the sequencing experiment will continue. However, sequencing at an incorrect temperature may lead to a decrease in throughput and lower q-scores. Please adjust the location of the sequencing device to ensure that it is placed at room temperature with good ventilation, then re-start the process in MinKNOW. Please refer to this link for more information on MinION temperature control. -
Guppy – no input .fast5 was found or basecalled
Observation Possible cause Comments and actions No input .fast5 was found or basecalled input_path did not point to the .fast5 file location The --input_path has to be followed by the full file path to the .fast5 files to be basecalled, and the location has to be accessible either locally or remotely through SSH. No input .fast5 was found or basecalled The .fast5 files were in a subfolder at the input_path location To allow Guppy to look into subfolders, add the --recursive flag to the command -
Guppy – no Pass or Fail folders were generated after basecalling
Observation Possible cause Comments and actions No Pass or Fail folders were generated after basecalling The --qscore_filtering flag was not included in the command The --qscore_filtering flag enables filtering of reads into Pass and Fail folders inside the output folder, based on their strand q-score. When performing live basecalling in MinKNOW, a q-score of 7 (corresponding to a basecall accuracy of ~80%) is used to separate reads into Pass and Fail folders. -
Guppy – unusually slow processing on a GPU computer
Observation Possible cause Comments and actions Unusually slow processing on a GPU computer The --device flag wasn't included in the command The --device flag specifies a GPU device to use for accelerate basecalling. If not included in the command, GPU will not be used. GPUs are counted from zero. An example is --device cuda:0 cuda:1, when 2 GPUs are specified to use by the Guppy command.
 In the image above, Sample 1 is of high molecular weight, whereas Sample 2 has been fragmented.
In the image above, Sample 1 is of high molecular weight, whereas Sample 2 has been fragmented. The pore activity plot above shows an increasing proportion of "unavailable" pores over time.
The pore activity plot above shows an increasing proportion of "unavailable" pores over time.Become a full member
Purchase a MinION Starter Pack from Avantor to get full community access and benefit from:
- News - hear about the latest product updates
- Posts - interact with thousands of nanopore users from around the globe
- Software - download the latest sequencing and analysis software
Already have a Nanopore Community account?
Log in hereNeed more help?
Request a call with our experts for detailed advice on implementing nanopore sequencing.
Request a callInterested in microbiology?
Visit our microbial sequencing spotlight page on vwr.com.
Microbial sequencing